 |
| 丂僩僢僾儁乕僕 > 儈僩僐儞僪儕傾偵嬊嵼壔偡傞夁巁壔暔姶庴惈寀岝僾儘乕僽 |
 |
||||||||||||||||||
儈僩僐儞僪儕傾偵嬊嵼壔偡傞夁巁壔暔姶庴惈寀岝僾儘乕僽
Abstract 丂Mitochondria are functionally important subcellular organelles fou丂our life. Reactive oxygen species 乮ROS乯 are produced without interruption in mitochondria. Some enzyme can be quenching of ROS in normal cells. Once the redox balance has broken down, ROS produced in mitochondria turn into intrinsic weapon. Thus, mitochondria are major source of ROS in mammalian cells and are major target of oxidative damage. Types of damages caused by ROS to mitochondrial components include lipid peroxidation, protein oxidation, and mitochondrial DNA mutation. In order to detect oxidative damage in living cells, some fluorescence probes have been developed. A new type of fluorescence probe, diphenylpyrenylphosphine-conjugated alkyltriphenylphosphonium iodide 乮MitoDPPP乯 which was accumulated in mitochondria, has been synthesized. This probe was detected peroxide in living cell. Comparison of the reactivity toward several peroxide indicated that the probe was existed in mitochondrial membrane. Using this probe, generation of peroxide in mitochondria of living cell was successfully visualized.
1丏 偼偠傔偵丂妶惈巁慺庬乮ROS乯偲偼丄僗乕僷乕僆僉僔僪傾僯僆儞儔僕僇儖 乮O2丒亅乯丄僸僪儘僉僔儖儔僕僇儖乮OH丒)丄堦廳崁巁慺乮1O2乯偍傛傃夁巁壔悈慺 H2O2 偺憤徧偱惗暔偺惗懚偵昁恵偺僄僱儖僊乕妉摼婡擻偦偺傕偺偵娷傑傟傞巁壔揑壔妛場巕乮儔僕僇儖乯偁傞偄偼壛楊丄巼奜慄丒曻幩慄丄戝婥墭愼丄栻暔偺挿婜暈梡丄墝憪偺桳奞惉暘偺媧擖側偳偵傛傝惗偠傞丅偦傟傜偼丄崅斀墳妶惈偱帀幙夁巁壔丄堚揱巕偺懝彎傗僞儞僷僋幙偺巁壔摍傪堷偒婲偙偟丄摦柆峝壔丄傾儖僣僴僀儅乕昦側偳偺幘昦偺堷偒嬥偵側傞丅崅楊壔偑壛懍偡傞変偑崙偵偍偄偰妶惈巁慺傗偦傟傜偺壔妛庬偵傛傝堷偒婲偙偝傟傞惗懱撪偱偺巁壔傪暘愅偡傞庤抜傪峔抸偟丄偦傟傜偺暘巕昦懺夝愅傪怺壔偡傞偙偲偑媫柋偱偁傞丅 1丏惗懱撪偱偺妶惈巁慺庬敪惗婡峔丂惗懱撪偱偺僄僱儖僊乕惗惉婡峔偵廳梫側栶妱傪壥偨偟偰偄傞偺偑儈僩僐儞僪儕傾撪偺揹巕揱払宯偱偁傝丄偦傟傜偼儈僩僐儞僪儕傾撪枌偵懚嵼偡傞僞儞僷僋幙暋崌懱偱峔惉偝傟偰偄傞丅偦偺暋崌懱傪捠偟偰丄儈僩僐儞僪儕傾枌娫僗儁乕僗偵僾儘僩儞傪曻弌偟丄偦偺僾儘僩儞傪嬱摦椡偲偟偰 ATP 傪崌惉偟偰偄傞丅嵶朎撪偵庢傝崬傑傟偨巁慺暘巕偺懡偔偼儈僩僐儞僪儕傾撪偵塣偽傟丄偦偙偱揹巕傪庴偗搉偟偝傟傞偙偲偱妶惈巁慺庬傊偲曄姺偝傟傞乮恾 1乯1乯 丅  偡側傢偪儈僩僐儞僪儕傾偼丄偙偺僄僱儖僊乕惗惉夁掱乮揹巕揱払宯乯偱丄條乆側妶惈巁慺庬傪惗惉偟懕偗傞惗懱撪偺妶惈巁慺庬敪惗尮偲側偭偰偄傞丅惗惉偟偨妶惈巁慺庬偼丄僗乕僷乕僆僉僔僪僕僗儉僞乕僛乮SOD乯丄僇僞儔乕僛丄僌儖僞僠僆儞儁儖僆僉僔僟乕僛側偳丄條乆側峺慺偑偦傟傜傪彍嫀偡傞偙偲偱嵶朎偺塩傒偼惓忢偵曐偨傟偰偄傞2乯丅偟偐偟側偑傜丄傂偲偨傃偙偺敪惗偲彍嫀婡峔偺僶儔儞僗偑曵傟傞偲丄儈僩僐儞僪儕傾偱敪惗偟偨妶惈巁慺庬偼丄峏側傞嫮椡側巁壔椡傪傕偮壔妛庬傪惗傒弌偡 3- 5乯 丅偙偺夁掱偱敪惗偡傞 OH丒偼丄僞儞僷僋幙丄帀幙丄DNA 側偳傪巁壔偟丄惗懱偵巁壔揑懝彎乮巁壔僗僩儗僗乯傪梌偊丄僈儞丄擼懖拞丄傾儖僣僴僀儅乕昦側偳偺堷偒嬥偲側傞6乯 丅懡偔偺妶惈巁慺庬偼儈僩僐儞僪儕傾偱敪惗偡傞偙偲偐傜丄儈僩僐儞僪儕傾偼惗懱撪偱嵟傕巁壔僗僩儗僗傪庴偗傗偡偄嵶朎撪旝彫婍姱偱偁傞偲偄偊傞7-9乯 丅  丂儈僩僐儞僪儕傾撪偱偺巁壔僗僩儗僗偼枌拞偵懚嵼偡傞晄朞榓帀幙偺楢嵔揑帀幙夁巁壔斀墳偲偟偰尰傟傞(恾 2乯丅偙偺斀墳偼丄儈僩僐儞僪儕傾枌傪峔惉偡傞晄朞榓帀朾巁偑丄 OH丒偵傛傝巁壔傪庴偗帀幙儔僕僇儖傪宍惉偟丄峏偵巁壔傪庴偗帀幙儁儖僆僉僔僪儔僕僇儖傪宱偰丄帀幙夁巁壔暔傪梌偊傞斀墳偱偁傞丅晄朞榓帀朾巁偑巁壔傪庴偗傞偲丄楢嵔揑偵斀墳偑恑峴偟丄帀幙夁巁壔暔傪拁愊偝偣傞丅偙偺帀幙夁巁壔暔偺拁愊傕傑偨丄榁壔丄摦柆峝壔傗僈儞側偳偺昦婥傪堷偒婲偙偡偙偲偐傜丄巁壔僗僩儗僗偺巜昗偲側傞10乯 丅偙偺帀幙夁巁壔偺専弌偵偼丄廬棃儓僂慺揌掕朄傗僠僆僶儖價僣乕儖巁乮TBARS乯朄偑梡偄傜傟偰偄偨偑丄偙傟傜偺應掕朄偼攋夡揑暘愅偱偁傝惗嵶朎拞偱偺娤應偱偼側偄丅傑偨 TBARS 朄偼丄帋椏拞偺儅儘儞僕傾儖僨僸僪傪専弌偡傞應掕朄偱丄偙傟偼帀幙夁巁壔暔偺擇師揑暔幙偱偁傞偨傔丄慖戰揑偵帀幙夁巁壔暔傪専弌偟偰偄傞傢偗偱偼側偄11乯 丅 2丏嵶朎枌摟夁惈寀岝僾儘乕僽丂嵶朎枌傪摟夁偟嵶朎撪偵摫擖偡傞偙偲偑偱偒傞寀岝僾儘乕僽偑偄偔偮偐曬崘偝傟偰偄傞丅僕僋儘儘僼儖僆儗僙僀儞僕傾僙僥乕僩乮DCF-DA乯傗 Peroxyfluor-1乮PF1乯12乯 偼嵶朎枌傪斾妑揑梕堈偵摟夁偟嵶朎幙偵廤愊偝傟傞乮恾 3乯丅摿偵 DCF-DA 偼丄傾僙僠儖婎偑嵶朎撪偺壛悈暘夝峺慺偵傛傝壛悈暘夝傪庴偗丄嵶朎撪懾棷惈偑崅偔側傞丅偟偐偟丄偙偺寀岝僾儘乕僽偼惗嵶朎拞偱嵶朎幙偵嶻惗偝傟傞妶惈巁慺庬傗崅妶惈儔僕僇儖庬傪曔懆偡傞偑丄壔妛庬傊偺慖戰惈傪帩偨側偄丅 PF1 偼丄僾儘乕僽帺恎偵棫懱揑偐偝崅偝傪帩偨偣傞偙偲偱彫偝側壔妛庬偵偺傒斀墳偡傞傛偆偵慖戰惈傪帩偨偣偰偄傞丅偟偐偟側偑傜丄帀幙夁巁壔暔偺傛偆偵棫懱揑偵偐偝崅偄壔妛庬偲偼斀墳偣偢丄偦偺嶻惗傪嵶朎拞偱専弌偡傞偙偲偼崲擄偱偁傞丅偙偺傛偆偵偐偝崅偄壔妛庬偵偺傒斀墳偡傞傛偆側慖戰惈傪帩偨偣傞偙偲偼寀岝僾儘乕僽偺暘巕愝寁偩偗偱偼摜攋偡傞偙偲偑偱偒側偄丅  3丏嵶朎撪旝彫婍姱嬊嵼壔寀岝僾儘乕僽丂愭偵傕婰偟偨傛偆偵丄儈僩僐儞僪儕傾偼丄妶惈巁慺庬嶻惗偲枾愙側娭學傪傕偮偙偲偐傜丄儈僩僐儞僪儕傾偵摿堎揑偵嬊嵼壔偡傞寀岝僾儘乕僽偺奐敪傕恑傫偱偄傞丅恾 4偵帵偡僕僸僪儘儘乕僟儈儞 123 傗 MitoAR 偼儘乕僟儈儞傪婎杮崪奿偵傕偮妶惈巁慺庬姶庴惈寀岝僾儘乕僽偱偁傞丅儈僩僐儞僪儕傾偼丄 ATP 崌惉偺嵺偵枌娫僗儁乕僗偵僾儘僩儞傪拁愊偝偣傞偨傔丄撪枌偲奜枌偲偺娫偵揹埵嵎偑惗偠丄偦偺揹埵嵎偼嵶朎宍幙枌傛傝傕崅偄丅偦偺摿幙偵墳摎偟偰儘乕僟儈儞偼儈僩僐儞僪儕傾偵嬊嵼壔偡傞丅僕僸僪儘儘乕僟儈儞 123 傗 MitoAR 偼嵶朎撪旝彫婍姱偵嬊嵼壔偡傞摿惈偼帩偮傕偺偺帀幙夁巁壔暔偵懳偡傞慖戰惈偑側偄13乯 丅僩儕僼僃僯儖儂僗儂僯僂儉墫偺傛偆側帀梟惈僇僠僆儞偼嵶朎撪傊庢傝崬傑傟傞偲丄揹埵嵎偑崅偄儈僩僐儞僪儕傾傊嬊嵼壔偡傞偙偲偑嬤擭 Murphy 傜偵傛傝曬崘偝傟偰偄傞14-17乯 丅偦偺僩儕僼僃僯儖儂僗儂僯僂儉墫傪 PF1 偵寢崌偝偣偨 MitoPF1 偑夁巁壔悈慺摿堎揑側姶庴惈傪傕偮寀岝僾儘乕僽偲偟偰奐敪偝傟偨丅 MitoPF1 偼丄PF1 偺摿惈傪偦偺傑傑庴偗宲偄偩儈僩僐儞僪儕傾偵嬊嵼壔偡傞寀岝僾儘乕僽偱丄棫懱揑偵偐偝崅偄妶惈庬偲偺斀墳惈偵朢偟偄丅偙偺傛偆偵嵶朎撪偱 OH丒丄O2丒亅丄 H2O2 側偳丄懡偔偺妶惈巁慺庬偑懚嵼偡傞拞丄儈僩僐儞僪儕傾撪帀幙夁巁壔暔傪慖戰揑偵幆暿偟壜帇壔偡傞傛偆側寀岝僾儘乕僽偼丄儈僩僐儞僪儕傾撪偱偺巁壔僗僩儗僗偺搙崌傪悇偟検傞偨傔偵昁梫偲側傞丅  4丏僩儕傾儕乕儖儂僗僼傿儞傪梡偄偨夁巁壔暔曔懆嵻丂3 壙儕儞壔崌暔偱偁傞僩儕傾儕乕儖儂僗僼傿儞傪寀岝僾儘乕僽偲偟偰棙梡偟偨曬崘椺偑偄偔偮偐偁傞乮恾 5乯乯18, 19乯 丅寀岝敪岝偡傞尨巕抍偱偁傞僺儗儞傗儁儕儗儞傪抲姺婎偲偟偰傕偮僕僼僃僯儖僺儗僯儖儂僗僼傿儞乮DPPP乯傗 3- 儁儕儗僯儖僕僼僃僯儖儂僗僼傿儞乮3- PeDPP乯偼娨尦忬懺偱偁傞儂僗僼傿儞偱偼儕儞尨巕忋偺屒棫揹巕懳傪夘偟偨暘巕撪岝揹巕堏摦斀墳偵傛傝寀岝抍傪徚岝偡傞丅  偲偙傠偑傂偲偨傃巁壔傪庴偗儂僗僼傿儞僆僉僔僪偵側傞偲偦偺徚岝偼夝徚偝傟寀岝抍偺嫮偄寀岝敪岝偑尒傜傟傞傛偆偵側傞丅偡側傢偪丄夁巁壔暔偵傛傝巁壔偝傟傞偙偲偱丄憡摉偡傞儂僗僼傿儞僆僉僔僪偲側傝丄寀岝嫮搙偑憹戝偡傞摿挜傪 DPPP 傗 3- PeDPP 偼傕偮乮恾 6乯丅偙偺傛偆側僩儕傾儕乕儖儂僗僼傿儞桿摫懱偼丄帀梟惈偑崅偔丄嵶朎撪摫擖偑崲擄偱偁傞丅嬤擭丄Soh 傜偵傛傝曬崘偝傟偨 Spy-LHP 偼帀梟惈偺崅偝傪棙梡偟丄偝傜偵傾儖僉儖嵔傪摫擖偡傞偙偲偱嵶朎宍幙枌偵嬊嵼壔偡傞夁巁壔暔曔懆寀岝僾儘乕僽偵摿壔偟偨桪傟偨寀岝僾儘乕僽偱偁傞偲偄偊傞20乯 丅  5丏儈僩僐儞僪儕傾偵嬊嵼壔偡傞帀幙夁巁壔暔姶庴惈寀岝僾儘乕僽丂儈僩僐儞僪儕傾偵嬊嵼壔偟夁巁壔暔傪曔懆偡傞寀岝僾儘乕僽偲偟偰丄変乆偼僩儕僼僃僯儖儂僗儂僯僂儉墫傪 DPPP 偵寢崌偝偣偨 MitoDPPP 傪奐敪偟偨乮恾 7乯丅 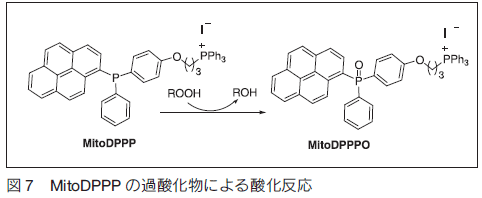 偙偺寀岝僾儘乕僽偼丄351 nm 偺岝偱椼婲偡傞偲 380 nm 偵寀岝敪岝傪帵偟丄巁壔偝傟偰 MitoDPPPO 偵側傞偲岝桿婲揹巕堏摦斀墳偵傛傞徚岝偑夝徚偝傟丄偦偺寀岝嫮搙偼栺 35 攞憹戝偡傞丅傑偨丄 MitoDPPP 偼丄梕堈偵嵶朎撪摫擖偝傟丄儈僩僐儞僪儕傾偵嬊嵼壔偡傞丅Methyl linoleate hydroperoxide乮MeLOOH乯丄Cumene hydroperoxide乮CumOOH乯丄tert-Butyl hydroperoxide乮t-BHP乯偍傛傃夁巁壔悈慺偲偺斀墳惈傪専摙偟偨偲偙傠丄嬒堦梟攠拞偱偼庒姳偺堘偄偼尒傜傟傞偑丄偳偺夁巁壔暔偵傛偭偰傕堦條偵巁壔偝傟夁巁壔悈慺偲偺斀墳偵偍偄偰傕懠偺帀梟惈偺夁巁壔暔偲傎傏摨摍偺斀墳惈偑尒傜傟偨乮恾 8乯丅  丂堦曽丄儕億僜乕儉拞偵偍偄偰偼丄帀梟惈偺夁巁壔暔傪梡偄偨斀墳偵斾傋傞偲丄夁巁壔悈慺偺偦傟偼抶偔側傞偙偲偑柧傜偐偲側偭偨乮恾 9乯丅  偝傜偵僸僩娞偑傫桼棃偺 HepG2 嵶朎拞偱庬乆偺夁巁壔暔偲偺斀墳惈傪斾妑偡傞偲丄帀梟惈偺夁巁壔暔偵傛偭偰懍傗偐偵巁壔傪庴偗傞偑丄夁巁壔悈慺偱偺巁壔偼嬌傔偰抶偄偙偲偑柧傜偐偲側偭偨乮恾 10乯丅 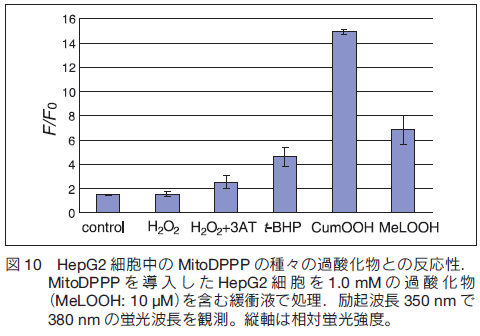
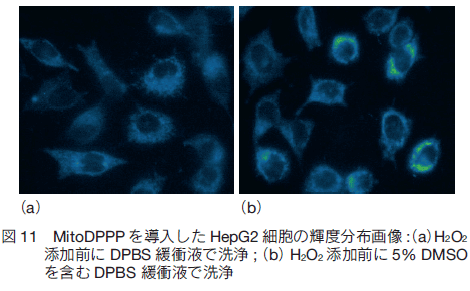
丂偟偐偟側偑傜丄偙偺寀岝僾儘乕僽偼丄寀岝抍偲偟偰僺儗儞婎傪梡偄偰偄傞偙偲偐傜丄嫟嶨帺壠寀岝偺塭嬁偲偦偺椼婲岝偺嵶朎傊偺岝撆惈偑婋湝偝傟傞偨傔寀岝尠旝嬀偺楢懕應掕偵偼揔偟偰偄側偄丅偙偺壽戣傪崕暈偡傞偵偼丄寀岝攇挿傪挿攇挿懁偵桳偡傞寀岝僾儘乕僽偺奐敪偑媮傔傜傟傞丅寀岝丄椼婲攇挿偲傕僺儗儞婎傛傝傕挿攇挿懁偵桳偡傞儁儕儗僯儖婎偵姺偊偨寀岝僾儘乕僽 MitoPeDPP 偼偙偺壽戣傪崕暈偟偨僾儘乕僽偱偁傞乮恾 12乯26乯 丅  偍傢傝偵丂崱夞丄嵶朎撪旝彫婍姱偵嬊嵼壔偟丄嵶朎撪偺僔僌僫儖暔幙傪専弌偡傞寀岝僾儘乕僽傪偄偔偮偐徯夘偟偨丅晹埵摿堎揑偐偮僔僌僫儖摿堎揑側庬乆偺寀岝僾儘乕僽傪暋悢慻傒崌傢偣傞偙偲偱嵶朎撪偺旝彫婍姱嬤朤偵偍偗傞僔僌僫儖暔幙偺嬊強揑側摦懺傪娤應偱偒傞偙偲偐傜丄崱傑偱尒傞偙偲偺偱偒側偐偭偨帠幚偺敪尒偵偮側偑傞偱偁傠偆丅傑偨丄偙傟傜偺僾儘乕僽傪棙梡偟偨峈巁壔嵻偺扵嶕丒奐敪偵傕婜懸偑傕偰傞丅 |
||||||||||||||||||
|
||||||||||||||||||

| Copyright(c) 1996-2015 DOJINDO LABORATORIES,丂ALL Rights Reserved. |