


- �@�@�@�@�@

���E�̃X�g���X�����ƃS���W�̃X�g���X����
ER stress response and Golgi stress response
 |
�g�c�@�G�Y ���Ɍ�����w ��w�@�������w������ ���� |
 |
���X�@�j�ލ] ���Ɍ�����w ��w�@�������w������ ���� |
Abstract
�@The endoplasmic reticulum (ER) and the Golgi apparatus are organelles where secretory and membrane proteins are correctly folded and post-translationally modified, respectively. When cells increase synthesis of secretory and membrane proteins, the capacity of the ER and the Golgi becomes insufficient (ER stress and Golgi stress), resulting in stress-induced cell death. To cope with these stresses, eukaryotic cells activate the homeostatic mechanisms called the ER stress response as well as the Golgi stress response, and increase the capacity of these organelles. In this review article, we described basics of the ER stress response and the Golgi stress response.
1. �͂��߂�
�@�זE�̒��ɂ͏��E�̂�S���W�̂ȂǗl�X�ȍזE���튯�����݂��邪�A�����זE��ł���ǂ̍זE�ł����ꂼ��̍זE���튯�̑��ݗʂ͂قڈ��ł���B�זE�����čזE���튯�̗ʂ������Ɍ����Ă��A�₪�čזE���튯�̗ʂ� 2 �{�ɑ����āA�ʂ����ɂȂ�悤�ɒ��߂���Ă���悤�Ɍ�����B�זE����ɂ���čזE���珬�E�̂��Ȃ��Ȃ��Ă��܂�����A�זE���S���W�̂��炯�ɂȂ��Ă��܂����Ƃ͂Ȃ��B�זE�ɊႪ���Ă���킯�ł��Ȃ��̂ɁA�ǂ�����Ē��߂��Ă���̂��A�ƂĂ��s�v�c�ł���B����A�R�̎Y���זE�Ȃǂ̂悤�Ƀ^���p�N���̕��傪����ȍזE�ł́A����^���p�N���̍����Ɖ��H���s�����E�̂�S���W�̂����������B���Ă���B�זE���튯�̗ʂ́A�זE�̎��v�ɉ����Č����ɒ��߂���Ă���悤�Ɍ�����B�����l����A�זE�����Ă��זE���튯�̗ʂ����ɕۂ���邱�Ƃ��������t���B���̂悤�ȍזE���튯�̗ʓI���ߋ@�\�͍זE�������I�ɋ@�\���邽�߂ɕK�{�̋@�\�ł���A�זE�����w�̍��{�I�Ȗ���̈�ł��邪�A�����Ԋj�̗ʓI���ߋ@�\�i�זE�����j�̌����ȊO�́A�قƂ�njڂ݂��邱�Ƃ��Ȃ����� 1�j�B�{�����ł́A���E�̋y�уS���W�̗̂ʓI���ߋ@�\�ł��鏬�E�̃X�g���X�����A�S���W�̃X�g���X�����ɂ��ĊT������B
2. ���E�̃X�g���X����
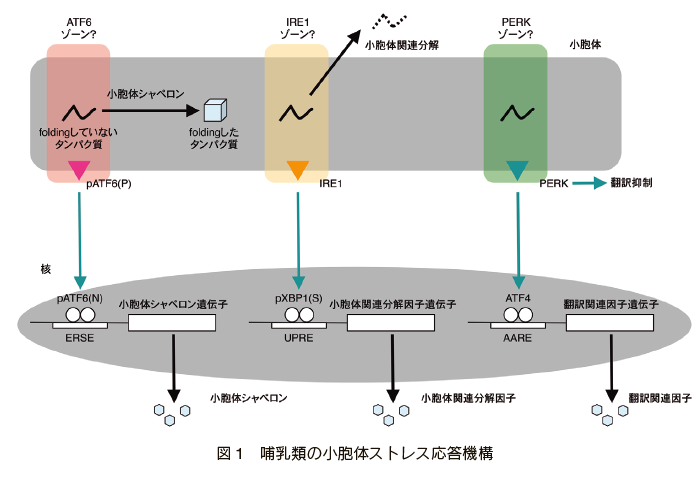
�@�X�a�r���m�i���s��w�j�́A���E�̗̂ʓI���ߋ@�\�ł��鏬�E�̃X�g���X�����̌����ɉʊ��ɒ��킵�A���̕��q�@�\�𖾂炩�ɂ��� 2, 3�j�B�M���ނ̏��E�̃X�g���X�����ɂ� ATF6 �o�H�� IRE1 �o�H�APERK �o�H�Ƃ��� 3 �̉����o�H�����݂��A���ꂼ��قȂ鏬�E�̂̋@�\�𐧌䂵�Ă���i�} 1�j�B���ꂼ��̉����o�H�́A �@���E�̂̋@�\���s�����Ă��邱�Ɓi���E�̃X�g���X�j�����m����Z���T�[�A �A�Z���T�[�ɂ���Ċ����������]�ʈ��q�A �B�]�ʈ��q����������G���n���T�[�z��A �C�]�ʂ��U�������W�I��`�q�� 4 ����Ȃ��Ă���B ATF6 �o�H�́A���E�̂ł̃^���p�N���t�H�[���f�B���O�\�͂�����o�H�ł���B�Z���T�[���q�� pATF6(P�j�́A�ʏ�͏��E�̖���ɑ��݂��Ă���B�t�H�[���f�B���O�\�͂��s�����ăt�H�[���f�B���O���������Ă��Ȃ��ُ�^���p�N�������E�̂ɒ~�ς���Ɓi���E�̃X�g���X��ԁj�A pATF6(P�j�͏��E�A���ɂ���ăS���W�̂֗A������A�S���W�̂ɑ��݂��� 2 �̃v���e�A�[�[�iS1P ��S2P�j�ɂ���Đؒf����Đ�����]�ʈ��q pATF6(N�j���G���n���T�[�z�� ERSE �Ɍ������邱�ƂŁA���E�̃V���y������`�q�̓]�ʂ�U������BIRE1 �o�H�́A���E�̂ɒ~�ς����ُ�^���p�N��������@�\�i���E�̊֘A�����iERAD�j�j������o�H�ł���B���E�̖���ɑ��݂���Z���T�[���q IRE1 �͕��펞�ɂ͒P�ʑ̂ł��邪�A�ُ�^���p�N�������m����Ƒ��ʑ̉����Ċ��������AXBP1 �̑O��� mRNA ���זE���ŃX�v���C�V���O���邱�Ƃɂ���Đ��n�^ mRNA �w�ƕϊ�����B���̐��n�^ mRNA ����|�ꂽ�]�ʈ��q pXBP1(S�j���G���n���T�[�z�� UPRE �Ɍ������邱�Ƃɂ���� ERAD ���q��`�q�̓]�ʂ�U������B PERK �o�H�́A�ꎞ�I�Ȗ|��}�����s���ƂƂ��ɁA�|��֘A���q��זE���̗U�����s���o�H�ł���B�Z���T�[�ł��� PERK �͏��E�̖���ɑ��݂��AIRE1 �Ƃ悭�����@�\�ɂ���Ċ���������B���������� PERK �͖|��J�n���q�ł��� eIF2 �� α �T�u���j�b�g�������_�����邱�ƂŖ|����ꎞ�I�ɗ}�����A����ȏ㏬�E�̃X�g���X���������Ȃ��悤�ɂ��Ă���B���̊Ԃ� ATF6 �o�H�� IRE1 �o�H�ɂ���ď��E�̓��ُ̈�^���p�N�������������悢���A�ˑR�Ƃ��Ĉُ�^���p�N�����~�ς��Ă���Ƃ��ɂ� PERK �o�H�ɂ���ē]�ʈ��q ATF4 �̖|�U������AATF4 ���G���n���T�[�z�� AARE �Ɍ������邱�ƂōזE����U������]�ʈ��q CHOP �̔������U�������B
3. �S���W�̃X�g���X����
�@�g�c�G�Y���m�i���Ɍ�����w�j�́A�X�a�r���m�̌���������Ɨ�����ۂɁA���E�ɐ�삯�ăS���W�̗̂ʓI���ߋ@�\�ł���S���W�̃X�g���X�����̌������J�n���A���݂͍��X�،j�ލ]���m�i���Ɍ�����w�j�ƂƂ��ɉ�͂�i�߂Ă���B�S���W�̃X�g���X�����ɂ������̉����o�H�����݂��A�S���W�̂̈قȂ�@�\�����Ă��� 4�j�i�} 2�j�B
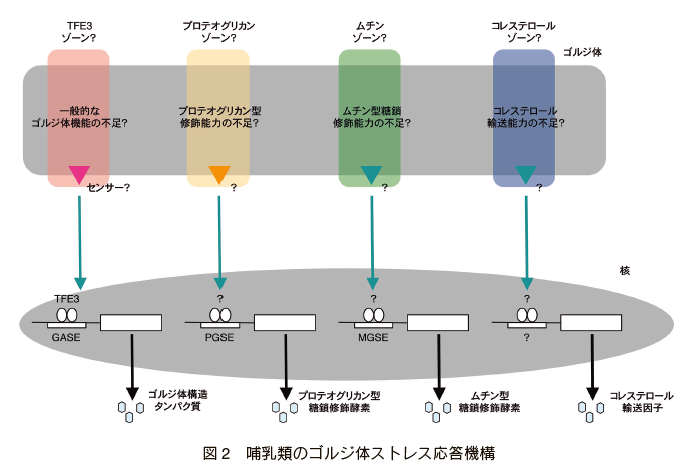
TFE3 �o�H�̓S���W�̂̈�ʓI�ȋ@�\������o�H�ł���A���̕W�I��`�q�ɂ̓S���W�̂̍\���^���p�N���ł��� GCP60 �⏬�E�A���Ɋ֗^���� GM130�AGiantin �Ȃǂ��܂܂��B�����̕W�I��`�q�̓]�ʂ𐧌䂷��G���n���T�[�z��� GASE�iGolgi stress response element�j�ƌĂ�A���̃R���Z���T�X�z��� ACGTGGC �ł��� 5�j�B TFE3 �o�H�𐧌䂷��]�ʈ��q�ɂ� bHLH-ZIP �^�]�ʈ��q�ł��� TFE3 6�j�� MLX 7�j������B���펞�ɂ� TFE3 �̓����_������čזE���ɗ��߂��Ă��邪�A�S���W�̃X�g���X���i�S���W�̂̈�ʓI�ȋ@�\���s���������j�ɂ͒E�����_������Ċj�ֈڍs���AGASE �Ɍ������邱�ƂŕW�I��`�q�̓]�ʂ�U������B MLX �� GASE �ɋ����I�Ɍ������邱�ƂŁATFE3 �̋@�\��}������]�ʗ}�����q���ƍl�����Ă���B
�@�v���e�I�O���J���o�H�́A�v���e�I�O���J���^�̓����C���\�͂����鉞���o�H�ł���B�v���e�I�O���J���͓�̏�����S�����q�ł��邪�A���̋@�\�ɂ̓v���e�I�O���J���^�̓����C��������߂ďd�v�ł���B�W�I��`�q�ɂ̓v���e�I�O���J���̓����C���y�f�◰�_���y�f�Ȃǂ̈�`�q�������܂܂�Ă���B�����̕W�I��`�q�̃v�����[�^�[��ɑ��݂���G���n���T�[�z�� PGSE�iproteoglycan-type Golgi stress response element�j���]�ʗU���𐧌䂵�Ă��邱�Ƃ��킩���Ă��邪 8�j�A�Z���T�[���q��]�ʈ��q�͂܂����m�ł���B��זE�Ȃǃv���e�I�O���J�����Y������זE�ł́A�v���e�I�O���J���o�H������������邱�ƂŁA�v���e�I�O���J���^�����C���\�͂����Ă���ƍl������B
�@���`���o�H�́A���`���^�̓����C���\�͂����鉞���o�H�ł���B���`���͑��ʂ̃��`���^�����C�����Ă���A�����̓������S�����`�����邱�Ƃɏd�v�ł���B�W�I��`�q�̓��`���^�����C���y�f�Q�̈�`�q�ł���A�G���n���T�[�z�� MGSE�imucin-type Golgi stress response element�j�����肳��Ă��� 9�j�B�Z���T�[�Ɠ]�ʈ��q�͖�����ł���B
�@�R���X�e���[���o�H�́A���E�̂���S���W�̂ւ̃R���X�e���[���̗A���\�͂����鉞���o�H�ł��� 10 �B�W�I��`�q�Ƃ��� OSBP2 �͓��肳��Ă��邪�A���̑��̐�����q�͓��肳��Ă��Ȃ��B
�@��L�̉����o�H�ȊO�ɂ��A���̌����҂��炢�����̉����o�H������Ă���B Jan Reiling ���m�iMIT�j�́ACREB3 �o�H�肵���B���ђʌ^�Z���T�[�ł��� CREB3 �������������� CREB3 ���ؒf����A������V������ CREB3 ���]�ʈ��q�Ƃ��ē����� ARF4 ��`�q�̓]�ʂ�U�����A���ʂƂ��ăA�|�g�[�V�X��U������ 11�j �B�{�c�M�ᔎ�m�i�ߋE��w�j�����肵�� HSP47 �o�H�́A�t�ɃS���W�̃X�g���X�ɂ��A�|�g�[�V�X��}������o�H�ł��� 12�j �B
5. ������
�@�ȏ�A���E�̃X�g���X�����ƃS���W�̃X�g���X�����ɂ��ĊT�������B���㏬�E�̃X�g���X�����ɂ��ẮA���̕��q�@�\�̏ڍׂȉ�͂��i�ނƂƂ��ɁA�̂ɂ����鏬�E�̃X�g���X�����̐����I�����̉�͂��i��ł������̂ƍl������B�S���W�̃X�g���X�����Ɋւ��ẮA������̐�����q�Q��ԗ��I�ɓ��肷��ƂƂ��ɁA�����̉����o�H�̃m�b�N�A�E�g�}�E�X�̕\���^����͂��邱�Ƃɂ���āA�S���W�̃X�g���X�����̑S�̑��𖾂炩�ɂ��Ă������Ƃ��̗v�ƍl���Ă���B
| [ ���҃v���t�B�[�� ] | |
| ���� | ���X�@�j�ލ]�iSASAKI Kanae�j |
|---|---|
| ���� | ���Ɍ�����w��w�@�������w������ ��678-1297�@���Ɍ��ԕ�S��S�� 3-2-1 |
| �o�g�w�Z | ������w��w�@�����_�w������ |
| �w�� | ���m�i�_�w�j |
| ��啪�� | �זE�����w�A�זE���튯�A�X�g���X���� |
| ���݂̌����e�[�} | �S���W�̃X�g���X���� |
| ���� | �g�c�@�G�Y�iYOSHIDA Hiderou�j |
|---|---|
| ���� | ���Ɍ�����w��w�@�������w������ ��678-1297�@���Ɍ��ԕ�S��S�� 3-2-1 |
| �o�g�w�Z | ���s��w��w�@���w������ |
| �w�� | ���m�i���w�j |
| ��啪�� | �זE�����w�A�זE���튯�A�X�g���X���� |
| ���݂̌����e�[�} | �S���W�̃X�g���X���� |