


セリン代謝酵素応答性蛍光プローブの開発
細胞は、外部から栄養(グルコース、アミノ酸、脂質など)を取り込み、さまざまな細胞内の代謝系を介して細胞の維持や増殖のためのエネルギー源や材料に変換し、利用している。また、代謝産物が細胞機能を厳密に制御していることが明らかとなり、細胞内代謝を理解することが重要とされている。近年、メタボローム解析や遺伝子操作などの技術の進歩により、細胞内の複雑な代謝経路が解明されている 1)。さらに、がん特異的な細胞内代謝は注目を浴び、新たな創薬標的候補として日々研究が進められている。
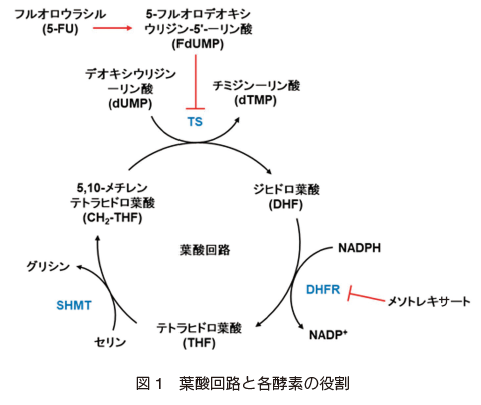
セリンヒドロキシメチル転移酵素(SHMT)は、アミノ酸の一種であるセリンをグリシンに変換する酵素であり、葉酸を介したアミノ酸や核酸合成に関わる One Carbon Metabolism の重要な酵素の一つである。この酵素は細胞増殖に寄与するため、がんやマラリアなど多くの疾病のバイオマーカーや治療薬の標的となっている。しかし、同じく One Carbon Metabolism に関わる重要なジヒドロ葉酸還元酵素(DHFR)やチミジル酸合成酵素(TS)阻害剤(メトトレキサート、5-FU)は、抗がん剤として長年使用されているのに対し(図 1) 2), 3),4)
、SHMT に関しては、未だ有用な阻害剤が開発されていない。
そこで本稿では、SHMT の酵素活性を簡便にモニタリングできる新規の蛍光プローブを用いた SHMT 阻害剤のスクリーニングについて紹介する 5)。
SHMT は上記のように、生化学的、医学的に重要であるにも拘らず、簡便かつ直接的に活性を測る手段が確立していない。その理由として、 SHMT の働きは、セリンからグリシンへの単純な反応であることに加え、酵素の基質結合部位が狭く設計可能な分子構造が限られていることが挙げられる。山東らは、 SHMT のセリン - グリシンの変換反応には葉酸が必要であるが、芳香族 β- ヒドロキシアミノ酸の変換には不要であることを見出した。さらに、後者の反応がレトロアルドール反応であること 6),7) を利用して、蛍光プローブβ-(p-dimethylaminonaphthyl)- serine(図 2)を開発した。
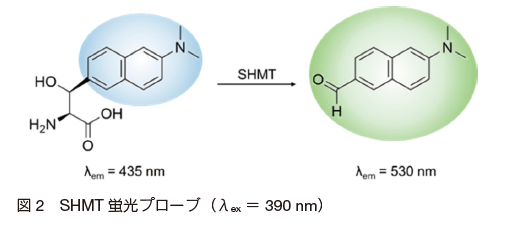
この蛍光プローブ自体は 435 nm の青色蛍光を発するが、SHMT との反応により 535 nm の緑色蛍光を有する p-dimethyamino-naphthaldehyde へと変換される(図 3)。
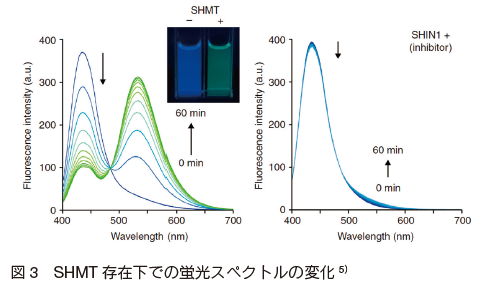
山東らはこの蛍光プローブを用いて、東京大学創薬機構が保有する約 21 万種類の化合物ライブラリーの中から、 SHMT 阻害剤の候補となる 2 種類の新規化合物を見出すことに成功している。得られた化合物は互いに類似性のないユニークな構造であり、有用な SHMT 阻害剤開発の手掛かりになると考えられる。さらに山東らは、 hydroxycoumarin 骨格の発蛍光型プローブや 19F-NMR 用のプローブを開発し、プローブ設計の柔軟さと蛍光プローブでは適用が困難な in vivo 実験への適用の可能性を示している。今回紹介した SHMT 応答性プローブは、 SHMT の作用機序解明や疾患の診断、および新たな抗がん剤や抗マラリア薬の開発への応用が期待される。
[参考文献]
-
1) Matsumoto, M. et al., Nat. Methods, 2017, 14, 251-258.
2) Locasale, J. W. et al., Nat. Rev. Cancer, 2013, 13, 572-583.
3) Jain, M. et al., Science, 2012, 336, 1040-1044.
4) Paone, A. et al., Cell Death Dis., 2014, 5, e1525.
5) Nonaka, H. et al., Nat. Commun., 2019, 10, 876.
6) Florio, R. et al., Biochim. Biophys. Acta., 2011, 1814, 1489-1496.
7) Webb, H. K. et al., J. Biol. Chem., 1995, 270, 17204-17209.