 |
| トップページ > 進化分子工学による人工酵素の設計と創出 |
 |
|||||||||||||||
進化分子工学による人工酵素の設計と創出
|
|||||||||||||||
|
||
|
要約
Advances in methods for conformational prediction, structural analysis and site-directed mutagenesis of proteins and peptides have contributed to the understanding of their structure and function. However, with the exception of a few successes, the generation of practical functional molecules solely by rational design remains a difficult challenge. The aim of our study is to investigate molecular design relying on evolutionary processes, called as “directed evolution”, to generate a novel class of biofunctional molecules. This evolutionary approach consists of three steps; 1) constructions of protein/peptide libraries based on structural information, 2) expressions of the libraries on phage particles, and 3) selections with investigator-imposed selective pressures. In this work, we study on generation of artificialbiocatalysts by immune system that uses the evolutionary processes to give receptor-like molecules. As a natural enzyme binds the transition state of the chemical reaction to lower the activation energy, immunization with a putative transition-state analog (TSA), with the expectation that the induced antigencombining site could be both geometrically and electronically complementary to the transition-state, provides catalytic antibodies. We have succeeded to generate antibodies catalyzing highly stereo and regio-specific hydrolyses with immunization of phosphonate transition-state analogs. Furthermore, to evolve catalytic antibodies toward higher catalytic activity, phage-displayed antibody (Fab) libraries were constructed to screen antibodies with optimized differential affinity for the transition state relative to the ground state. Recently, we have demonstrated a new strategy for generating catalytic antibodies, namely, by the generation of antigen-combining site that function as an apoprotein for binding functionalized components, called “holoabzyme”. Replacement of the functionalized components enables a single antibody to catalyze multiple reactions, β -elimination, decarboxylation and aldol reactions. Catalytic antibodies have great potential for generating novel catalysts as well as for providing opportunities to examine the evolutionary dynamics of enzymes.
キーワード:1) 抗体、 2) 抗体酵素、 3) 進化分子工学、 4) ファージ表層提示ライ
ブラリー、 |
||

1.はじめに
新しい機能を持つ人工タンパク質を自由自在に創り出すこと は、化学・生物工学の最終目標の1つである。この20 年の遺伝 子工学や構造生物学のめざましい進展は、タンパク質の大量調製 や部位特異的変異操作による修飾酵素の合成を可能にしている。 今や、タンパク質は、対応するDNAを合成するだけで、機械的 に合成できる。しかしながら、これまでの研究からわかるように、 水素結合などのネットワークを考慮して機能発現に必要なアミノ 酸を適切な部位に正確にならべることは難しい。研究者はDNA合成機の前で「どのような設計図を書いたらいいのだろうか」と 苦吟しているのが現状である。
それでは、生細胞のDNA上にあるすばらしいタンパク質の設
計図は、どのようにして書かれたのだろうか?いまのところ、こ
の設計図はダーウィン進化の過程でDNA上に間接的に書き込ま
れたと考えられている。すなわち、生細胞は、長い年月をかけて
突然変異を蓄積して分子多様性をもたらし、選別を繰り返して酵
素のような高度な触媒機能をもつタンパク質を獲得する。最近、
このような自然界における進化の過程「多様性の発生と選別」を
人為的にコントロールして、機能性タンパク質の創出を目指す進
化分子工学(自然進化と区別するためにコンビナトリアル・バイ
オエンジニアリングとも呼ばれる)が注目されている。多様な化
合物を一挙に合成する「コンビナトリアル・ケミストリー」が知
られているが、進化分子工学では、生物学的手法を組み合わせ、
人工的な生体触媒や生理活性分子を創出することを狙っている。
すなわち、免疫システム(抗体ライブラリー)やファージ表層提
示法(ペプチドやタンパク質ライブラリー)により、多様な分子
ライブラリーを効率的にスクリーニングし、目的とした機能を持
つ生体分子を獲得する手法である。本稿では、免疫システムから
生まれる抗体タンパク質を取り上げて、進化分子工学による人工
酵素の創出について紹介する。
2.免疫システム
免疫システムは、外から侵入してくる抗原に応答し、高い親和 性と特異性を持つ抗体タンパク質を産生する。侵入してくる抗原 は多種多様なので、それらに対応するためには特別な抗体産生の 仕掛けが必要になる。抗体タンパク質は、「多様性の発生、提示、 選別」の過程を経て創られている(図1)。すなわち、免疫シス テムは、生体が私たちに与えた進化分子工学であり、これを利用 することで目的とした触媒活性をもつ人工酵素を手に入れること ができる。
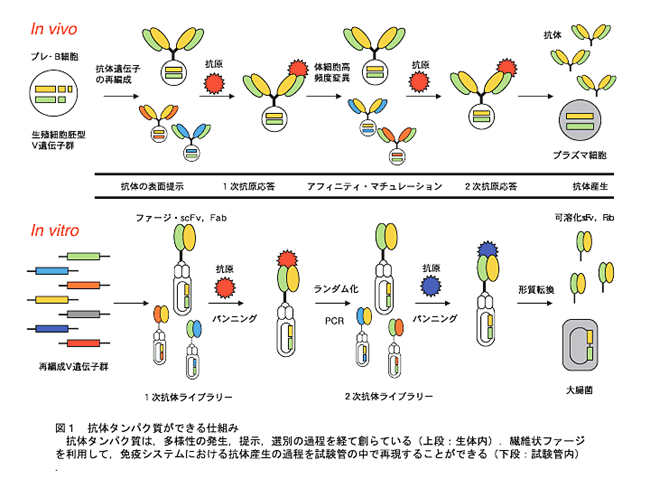
抗体タンパク質の多様性は、抗体の抗原結合部位をコ-ドする 遺伝子セグメント(H鎖:VH,D,JH遺伝子群、Lκ鎖:Vκ,Jκ遺 伝子群)の組み合わせ(コンビナトリアル)により生ずる。マ ウスの場合、VH遺伝子群は100個以上の遺伝子があり、 D遺伝子は約15個、JH遺伝子は5個存在する。また、L鎖を形成す るVκ遺伝子群は200~300個、Jκ遺伝子群は5個の遺伝子か らなる。これらの遺伝子が、B細胞が骨髄中の幹細胞から成熟B細胞に分化・成熟する過程で再編成される。このような仕組みで 生じた特異性の異なる抗体タンパク質(108種)は、B細胞表面 に提示され、外来抗原の侵入に備えている。第1免疫応答では、 抗原に親和性のある抗体を提示しているB細胞がプラズマ細胞 あるいは記憶細胞へと分化・増殖を始める。この増殖の過程で体 細胞高頻度変異が起こり、抗体の多様性がさらに増える。あるも のは体細胞高頻度変異の結果、親和性が低下し、あるものでは親 和性が向上する。そこで、第2・第3免疫に応答して、親和性の より高い抗体を提示しているB細胞が選択され、分化・増殖す る(この過程をアフィニティ-・マチュレ-ションと呼ぶ)。す なわち、免疫システムは、生体が私たちに与えた進化分子工学で あり、これを利用することで目的とした結合活性をもつ生体機能 分子を手に入れることができる。
3.テーラーメイド人工酵素:抗体酵素
抗体タンパク質を使って酵素の触媒機能を創出する試みがあ る。しかしながら、酵素の触媒機能を抗体で再現するには、抗体 の選択基準を工夫する必要がある。その1つは遷移状態アナログ に対する親和性を基準として抗体を選択する方法である。すなわ ち、酵素が化学反応の遷移状態と結合し安定化することによって 触媒機能を発揮しているように、遷移状態アナログに結合する抗 体タンパク質は、化学反応の遷移状態と結合し触媒機能を獲得す るようになる。このような触媒活性を持つ抗体を「抗体酵素」と 呼ぶ1,2)。酵素が何十億年という長い年月をかけて進化したのに 対し、抗体の進化は数週間で起こるので、実験室で測定可能な時 間内に分子進化の過程を再現して、テーラーメイドの生体触媒を 創ることができる。
3.1抗体酵素の原理
一般に化学反応の触媒現象は、エントロピー因子、酸-塩基触 媒、静電的効果などの触媒因子の組み合わせによって起こるもの と理解されている。これに対して酵素触媒反応の大きな特徴の一 つは、酵素が化学反応の遷移状態に結合し、それを安定化するこ とによって反応の活性化エネルギーを減少させ、化学反応を促進 していることである(図2)。抗体酵素の触媒作用も、酵素と同 様に反応の遷移状態を安定化するということに基づいている。す なわち、反応の遷移状態と立体的かつ電子的によく似ている化合 物「遷移状態アナログ」をハプテンとして免疫すると、得られた 抗体は反応の遷移状態と結合し、安定化することによって反応を 触媒する。ハプテンとは、それ自身では抗原性をもたない低分子 化合物で、キャリアータンパク質(ウシ血清アルブミンなど)と 結合させることにより抗原性を獲得する。
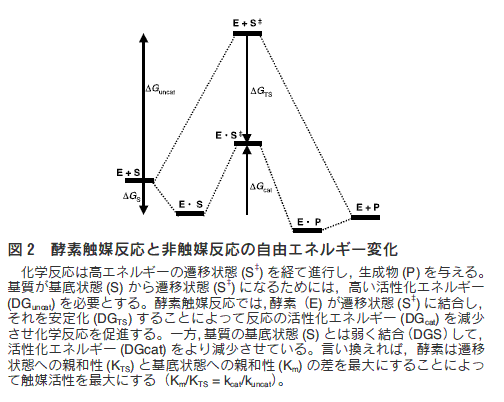
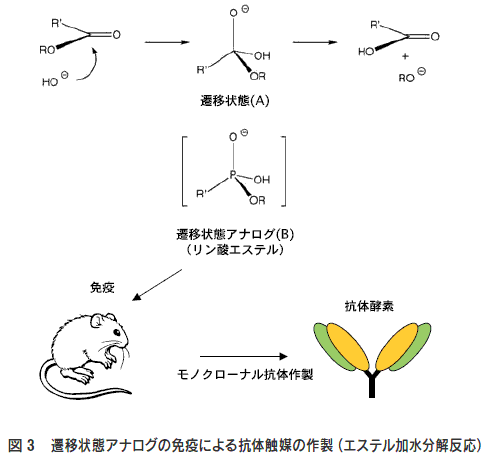
図3にエステル結合の加水分解反応の例を示した。エステルの 加水分解はカルボニル基の酸素原子上に負の電荷を持つ高エネル ギーな四面体遷移状態(A)を経て進行する。この四面体遷移状 態(A)に立体的かつ電子的に相補的な抗原結合部位を持つ抗体 は、反応の活性化自由エネルギーを減少させ、加水分解反応を触 媒する。現在、遷移状態アナログとしてリン酸エステル(B)が 使われており、これを免疫する事によってエステル加水分解を触 媒する抗体の作製が行われている3)。 これまでに筆者らは、独自の遷移状態アナログ設計により立体選択 的反応4-6) や位置選択的反応など 7-9)を触媒する抗体の作製に成功している。
3.2.抗体酵素によるプロドラッグ医薬品の活性化
免疫システムを利用して、プロドラッグ医薬品を特異的に活性 化する人工酵素を作製した10,11)。プロドラッグとは、医薬品の 物性の改善や毒性軽減を目的として化学修飾された化合物であ り、それ自身は薬理活性を示さないが、投与後生体の酵素やpH の変化により元の医薬品(親化合物)に変換されて薬理作用を発 揮するものである。もし、生体内の天然酵素に対して安定なプロ ドラッグをテーラーメイドの抗体酵素で活性化することができれ ば、病巣特異的なドラッグ・デリバリーが可能になる。そこで、 モデル医薬品として抗生物質であるクロラムフェニコール(1)を 取り上げ、そのプロドラッグであるエステル(2) を加水分解する 抗体を作製した(図4)11)。
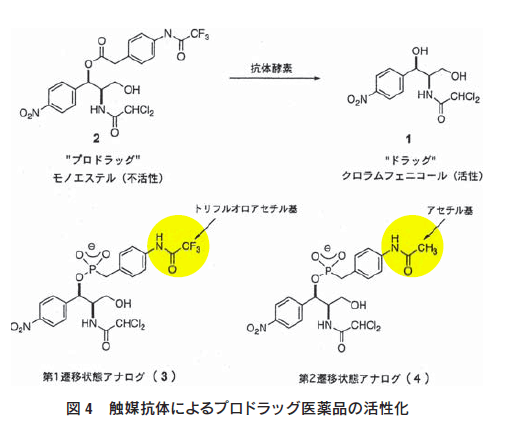
エステル(2)の加水分解反応の遷移状態アナログであるリン酸 エステル(3)を合成した。これをキャリアータンパク質(KLH) と縮合後、抗原としてマウスに免疫し、モノクローナル抗体を作 製してハプテンに結合活性を持つ12種類の抗体を得た。この抗 体について、エステル(2)の加水分解活性を検討したところ、6種類の抗体に顕著な触媒活性が観測された。もっとも活性の高 かった抗体(6D9)は、抗体のない場合に比べて反応を800倍以上加速した(表1)。
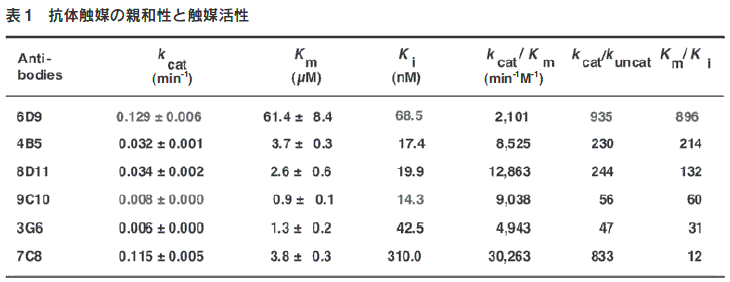
3.3.抗体酵素の触媒機構
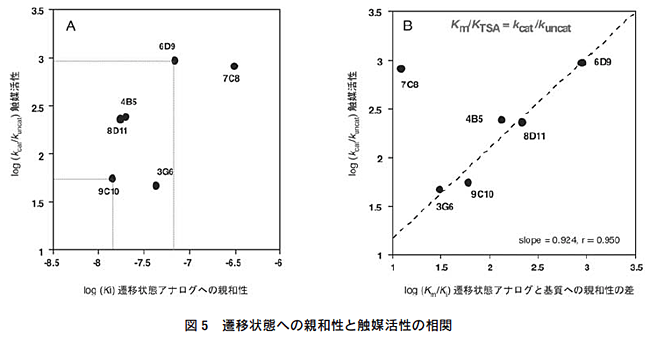
遷移状態アナログ (3) の免疫から得られた抗体の結合活性と触 媒活性を表1に示す。抗体酵素の反応速度論量を比較すること によって、遷移状態アナログ対して高い親和性を持つ抗体が、必 ずしも高い触媒活性を示すとは限らないということが判明した12)。最も親和性の高い抗体(抗体9C10:KTSA=0.014μM)は最も触媒活性(kcat=0.008min-1)が低く、また高活性の抗体6D9(kcat=0.129min-1)は中程度の親和性(KTSA=0.068μM)を示す(図5A)。一方、反応加速(kcat/kuncat)と基質の基底状態と遷移状態に対する結合活性の差(Km/KTSA)に相関性が観測された。抗体6D9をはじめとして8D11,4B5,9C10,3G6の5種の抗体では、kcat/kuncat値とKm/KTSA値がほぼ一致した(図5B)。 この解析から、触媒作用の発現するために重要なことは、遷移状態に対する強い結合と基底状態への弱い結合であり、親和性の差の大きい抗体が高い触媒活性を持つことが判明した。すなわち、抗体酵素に高い活性を付与するためには、遷移状態への結合だけでなく、遷移状態と基底状態との親和性の差が最大になるような抗原結合部位を構築する必要がある。
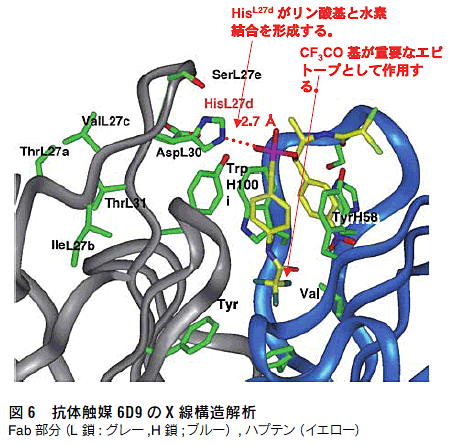
抗体酵素の触媒機構を明らかにするために、抗体6D9 のX線結晶解析を行った13)。抗体酵素6D9では図6に示すように、ハプテンの2個のベンゼン環は、Tyr(H58)とTrp(H100i)とσ-π相互作用で抗原結合部位の深い位置に取り込まれている。また、遷移状態を模倣しているホスホン酸エステル部の酸素原子は、抗原結合部位の入り口に位置するHis(L27d)と水素結合している。部位特異的変異操作によりHis(L27d)をAlaに置換した抗体は全く触媒活性を失う14)。以上の結果より、抗体酵素6D9においては、L鎖CDR-1領域のHis(L27d)が遷移状態のオキシアニオンと水素結合あるいは静電的相互作用し、遷移状態を安定化して触媒機能を発現している15,16)。
以上のように、一連の抗体酵素はハプテンの設計から期待される遷移状態の安定化を触媒因子として、その機能を発現している。 すなわち、抗体の触媒機能は、免疫系の多様性から生まれる偶発的なものではなく、合理的なハプテンの設計によって発現される ものといえる。
4.ファージ・ディスプレイ・システム
繊維状ファージは,大腸菌に感染する非常に単純なウイルスであり、遺伝子工学においてさまざまな用途に用いられている。こ のような繊維状ファージを利用することによって、免疫システムにおける抗体産生の過程を試験管の中で再現し、抗体タンパク質 の機能を改良することができる17)。
4.1.抗体の進化分子工学
ファージディスプレイを用いた抗体分子の選択は、基本的には、 図1に示したような生体内の免疫系の働きである「多様性の発 生」「提示」「選別」を模倣している。すなわち、1)多様性を持 つ抗体遺伝子を増幅する、2)ファージを抗体産生B細胞になぞ らえ提示する、3)目的とする機能で選択するの3段階をすべて 試験管内で行うことになる。さまざまな特異性を有する分子を分 子ごとに表面にディスプレイすることで、コンビナトリアルなラ イブラリーを作り出し、ディスプレイされた抗体タンパク質が持 つ機能に基づいて、新しい分子を選択することができるわけであ る。この手法により、実験動物の免疫化を経ることなく抗体分子 を手にすることが可能になっているほか、免疫化が困難で取得が 困難であった抗体の選択・調製が可能となり、さまざまな応用に 成功を収めている。ここでは、ファージディスプレイを利用した 抗体酵素の機能改変について述べる18,19,20)。なお、ファージディスプレイ法の詳細な手法については、参考文献に挙げた論文や他 の成書を参照していただきたい21)。
4.2.抗体酵素の高活性化
先に述べたプロドラッグ医薬品を特異的に活性化する抗体酵素 (6D9)の高活性化に際して、ファージ・ディスプレイ・システ ムを利用した19,20)。この方法により免疫法では取得することの できない高い活性を持つ抗体酵素の作製が可能になる。
酵素などの生体触媒の活性発現に重要なことは、遷移状態に対 する強い結合と基底状態への弱い結合である。先に述べたように、 抗体酵素の場合にも、高い活性を得るためには、遷移状態への結 合だけでなく、遷移状態と基底状態との親和性の差が最大になる ような抗原結合部位を構築する必要がある。ところが、抗体産生 のメカニズムを考えると、一度の免疫で2つの分子種(遷移状 態と基底状態)の情報をもつ抗体を作製することは不可能である。 そこで、抗体産生の過程をファージ抗体を用いて試験管内で再現 し、遷移状態と基底状態との親和性を差を最大にさせるように抗 体の分子認識を最適化した。この方法は、従来の免疫法とは異な り、抗体遺伝子に任意に変異を導入したり、また多種の抗原で何 度もスクリーニングしたりできることから、自由自在に抗原への 親和性や抗原特異性を変化させることができる。
遷移状態と基底状態との親和性の差を大きくするためには、ハ プテン分子全体よりも遷移状態を模しているリン酸エステル部分 (四面体構造)に強く結合する抗体を選択する必要である。そこで、 第1遷移状態アナログ(3)のトリフルオロアセチル基をアセチ ル基に変換した第2遷移状態アナログ(4)を合成し、これを使っ てファージ抗体ライブラリーのスクリーニングを行った(図4)。 遷移状態アナログ(3)のトリフルオロアセチル基は重要なエピ トープなので、アセチル基に変換することで抗体の親和性が弱め られる。こうすることで、一旦、抗原全体への親和性を下げ、リ ン酸エステルの四面体構造部分により強い結合を持つようになっ た抗体を選別する。
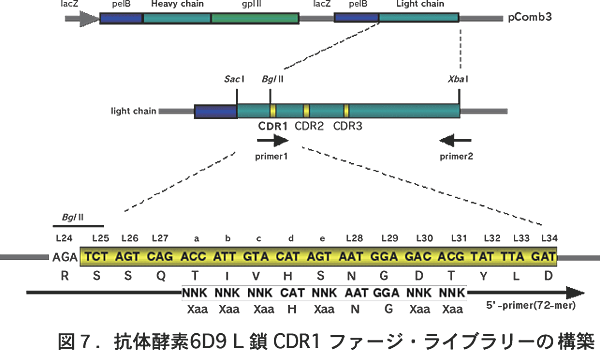
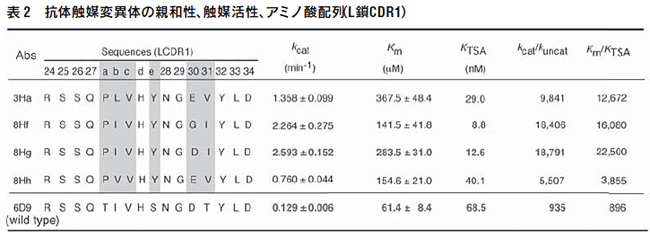
X 線構造解析や部位特異的変異実験から、抗体6D9 のL鎖 CDR1 のHis(L27d)が触媒残基として働いていることが判明し ている(図6)13,14)。そこで、活性中心のHis残基は固定し、そ の周辺のアミノ酸6残基に対してランダム変異を行い、抗体ライ ブラリーを作製した(図7)。そして、第2遷移状態アナログ(4) に対して、この抗体ライブラリーを8回パンニングし、ハプテン 結合性の抗体を選別した。
得られた変異体は野生型(6D9)に比べて高い活性を示し、変 異体(8Hg)では20倍以上強い触媒活性を獲得していた(表2)。 従来の免疫法から得られた抗体酵素が、無触媒反応に比べ50~1000倍程度の反応加速を示すのに対し、ファージ抗体ライブラリー からは反応加速が6000~20000倍の抗体酵素を得ることに 成功した。変異体の分子モデルを作成したところ、触媒残基His (L27d)のとなりにあるSer(L27e)がTyrに変わっていることが判 明した(図6)。すなわち、野生型ではHis(L27d)が遷移状態と 水素結合を形成し安定化させ、一方、変異体ではHis(L27d)と ともに新しく導入されたTyr(L27e)とで2つの水素結合を作り、 より強く遷移状態を安定化する。さらに、アミノ酸配列の比較に より、ファージディスプレイ法の有用性が示された。Tyr(L27e) はSerから変異したものであるが、Ser(AGT)からTyr(TAT, TAC)への変異には核酸レベルで隣り合った2つの変異が起こる ことが必要である。このような突然変異は抗体産生の過程(体細 胞変異による親和性成熟)では起こり難い。すなわち、得られた 高活性抗体酵素を従来の免疫法で得ることは困難であり、ファー ジディスプレイ法の有用性が示された。
5.抗体酵素が触媒する化学反応
抗体の特徴は、抗原に対して特異的に結合する高い分子認識で
ある。特に、抗体酵素の抗原結合部位は天然酵素と同様に「不斉
反応の場」となっているので、不斉反応への展開がもっとも期待
される(図8)。通常、この種の反応は、 メソ化合物のエナン
チオ場区別反応、
メソ化合物のエナン
チオ場区別反応、 プロキラル化合物のエナンチオ面区別反応、
プロキラル化合物のエナンチオ面区別反応、 エナンチオマー区別反応(速度論的光学分割)の3つに分類す
ることができる。エステル結合を加水分解する抗体酵素によって、
このようなエナンチオ区別反応が可能になる。
エナンチオマー区別反応(速度論的光学分割)の3つに分類す
ることができる。エステル結合を加水分解する抗体酵素によって、
このようなエナンチオ区別反応が可能になる。
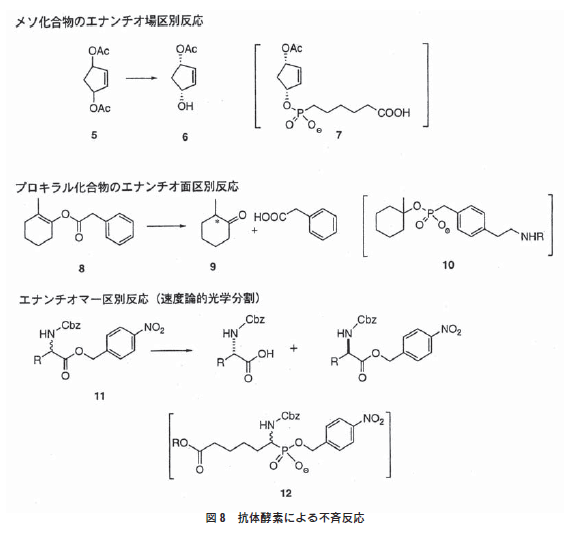
5.1.メソ化合物のエナンチオ場区別反応
メソ化合物(5)から光学活性アルコール(6)へ加水分解反応を 触媒する抗体が、ホスホン酸エステル(7)をハプテンとして作製 された22)。得られた抗体の触媒活性は、本反応を触媒するアセ チルコリンエステラーゼに比べてかなり低いものであったが、生 成したアルコール(6)は86%の光学純度を示す。
5.2.プロキラル化合物のエナンチオ面区別反応
ホスホン酸エステル(10)を免疫して得られる抗体酵素は、α位にメチル基をもつエノールエステル(8)の加水分解において、 プロトン化が片方の面から優先的に起こり光学活性のα-メチルケトン(9)を与える4)。
5.3.エナンチオマー区別反応(速度論的光学分割)
α-置換基を有するアミノ酸エステル誘導体(11)の両エナン チオマーのそれぞれに対して、エナンチオ選択的加水分解反応を 触媒する抗体がハプテン(12)の免疫によって作製される5)。ハプ テン自身はラセミ体だが、抗体を光学活性のアミノ酸エステルを 基質としてスクリーニングすることにより、L-体選択性抗体お よびD-体選択性抗体を得ることができる。これらの抗体はアラ ニン、ロイシン、ノルロイシン、メチオニン、フェニルアラニン、 バリン、フェニルグリシンや非天然アミノ酸の4-ヒドロキシフェ ニルグリシンに対しても触媒活性を示し、高い立体選択性と同時 に幅広い基質特異性がある。
5.4.位置選択的反応
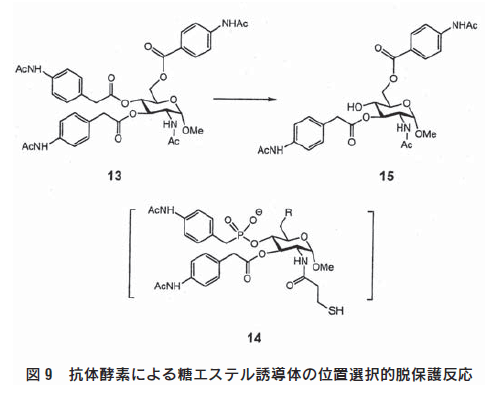
抗体酵素の厳密な分子認識は、分子内に多くの同一官能基を有 する糖類の位置及び立体特異的反応に適している。安価で容易に 入手できる糖類は、有機合成の不斉制御素子や不斉配位子の出発 原料として繁用されている。糖類を基質として合成を行うには、 任意の水酸基を他の水酸基と区別することが重要な問題となって くる。特に、糖鎖合成の場合には、特定の水酸基から結合をさせ ようとすると、水酸基の保護-脱保護の操作を繰返さなければな らず、結果的に目的物を低収率でしか得られないことが多い。こ のような糖基質中の水酸基の保護-脱保護にかかる労力を軽減で きれば、糖質関連化合物の合成をより効率的に行える。そこで、 糖基質の構造を認識して複数のエステル結合の1つを選択的に切断 する抗体酵素が作製された(図9)7)。
基質としてグルコサミンの3、4、6位の水酸基の4-トリフル オロアミノベンジルエステル基で保護したトリエステル(13)が 用いられた。この基質の4位エステル基だけを位置選択的に脱保 護するために、4位にホスホン酸エステル基をもつハプテン(14)の免疫により抗体が作製された。得られた抗体は、Michaelis-Menten型反応速度式に従って4位エステル基を加水分解し(kcat=0.182min-1,Km=6.6μM,kcatlkuncat=2700)、生成物(15)を与える。この抗体酵素は位置選択性とともに立体選択性を示し、4位の立体異性体であるガラクサミン誘導体には作用しない。
6.ホロ抗体酵素
これまでに多くの抗体が創られ多種多様な有機化学反応を触媒 することが示されている。これらの触媒抗体の多くは遷移状態 アナログの免疫によって作製されている。そこで、筆者らは、従 来の遷移状態アナログを免疫する手法とは全く異なる新しい触媒 抗体の作製法の開発を目指して、ホロ酵素型触媒抗体の分子設 計を検討した23)。ホロ酵素は、活性部位に低分子有機化合物のコファクターをもち、これを触媒基として化学反応を触媒する。そ こで、抗体をアポ酵素として利用し、基質分子に加えて、人工合 成コファクター分子に対する抗原結合部位の構築を行った。図10に示すように、反応性の異なる人工合成コファクター分子を 設計し、それらを入れ替えることにより、抗体が触媒できる反応 の種類や触媒機構を制御することを目指した。
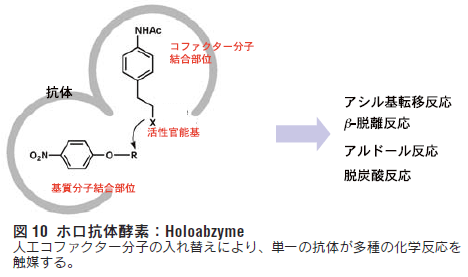
6.1.ホロ抗体酵素の作製
リン酸モノエステル(B)をマウスに免疫することにより作製 されたエステルの加水分解反応を触媒する抗体は、その抗原結合 部位に遷移状態を安定化するアミノ酸残基(ヒスチジン、チロシ ンなど)が(B)の有する負電荷により誘導されるため、エステ ルの加水分解反応の遷移状態に特異的に結合し、安定化すること によって反応を加速する。また、抗体はもともと非常に高い特異 性を有しているため、この抗体がエステルの加水分解以外の反応 つまり、遷移状態の全く異なる反応を触媒することは不可能であ る。これは、天然酵素であっても同様である。そこで、複数の有 機化学反応を高効率で触媒するホロ酵素型触媒抗体の開発を目指 し、図11に示すようなリン酸ジエステルハプテン(16)の設計を 行った。
ハプテン(16)は2つの特徴を持っている(p-ニトロフェニ
ルリン酸エステル部分は、アシル基転移反応の遷移状態を模擬す
ると同時に抗原結合部位に基質分子結合部位を与える、N-アセ
チルフェネチル部分は種々の人工コファクター分子結合部位を抗
原結合部位に構築する)。また、電荷を持たないリン酸ジエステル
ハプテンを免疫に用いることにより、アシル基転移反応の遷移状
態を強く安定化するようなアミノ酸残基が誘導されることに起因
する遷移状態の安定化を触媒因子とするというよりも、基質分子
および人工コファクター分子の2分子を幾何学的に適切な位置に
置くことによる近接効果を主要な触媒因子とする触媒抗体の作製
が期待される。これにより求核触媒、酸・塩基触媒能をもつ補酵
素あるいは人工コファクター分子を抗原結合部位に導入し、それ
らを入れ替えるという新しい概念を導入することにより、人工コ
ファクター分子により触媒できる反応の種類を制御可能な、天然
にはない機能性をもつ触媒抗体の開発につながると期待される。
基質分子としてエステル(17)を用い、人工コファクター分子 としてヒドロキシル基をもつ化合物(18)、アミノ基をもつ化合 物(19)、チオール基をもつ化合物(20)を添加すれば求核触媒と してアシル基転移反応を、基質としてβ-ハロケトン(23)を用い、 人工コファクター分子としてカルボキシル基をもつ化合物(24)を添加すれば塩基触媒としてβ-脱離反応を、人工コファクター 分子としてアミン分子(19)を添加すればエナミン機構を経るア ルドール反応および脱炭酸反応を触媒することが期待される(図11)。
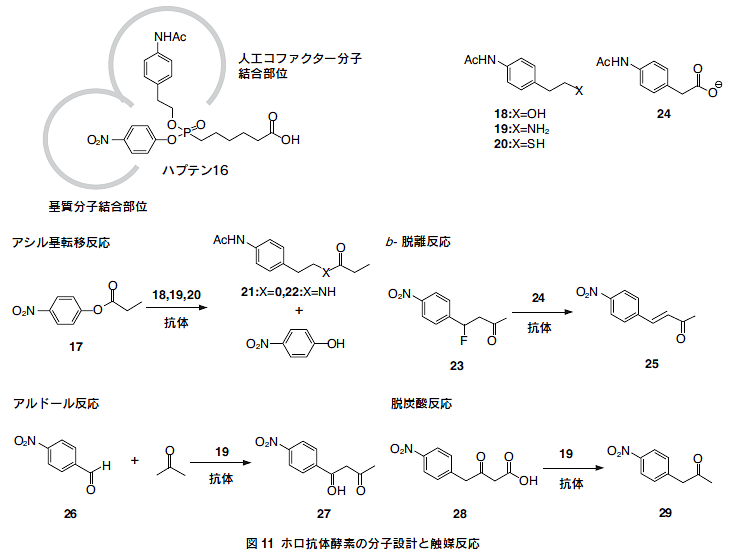
6.2.アシル基転移反応
ハプテン(16)を担体タンパク質であるスカシ貝ヘモシアニン (KLH)やウシ血清アルブミン(BSA)への縮合を行い、KLH-ハ プテン縮合体(KLH-1)をBalb/cマウスに免疫した。常法により、 ハプテン(16)を特異的に認識する50種類のモノクローナル抗体 が得られた。
次に、50種の抗体に関して活性スクリーニングを行った。ハ プテン(16)の設計からまず考えられるエステル(17)とアルコー ル(18)とのアシル転移反応を検討した結果、22種の抗体に顕著 な触媒活性を観測した。そこで最も活性の高かった2種の抗体 (25E2,27C1)について、詳細な反応速度論的解析を行った。人 工コファクター分子としてアルコール(18)、アミン(19)、チオー ル(20)のいずれを用いた場合にも高い触媒活性を示した。アル コール(18)を用いた場合198倍(25E2)、109倍(27C1)、アミン (19) を用いた場合1.4×104倍(25E2)、5.5×104倍(27C1)、チ オール(20)を用いた場合4.2×103倍(25E2)、3.0×103倍(27C1) の速度加速を示した。また、両抗体はハプテン(16)に対し非常に高い結合能を有し(25E2:Kd=27nM;27C1:Kd=8.7nM)、さ らに抗体の触媒するすべての触媒反応(アシル転移反応、β-脱 離反応、アルドール反応および脱炭酸反応)はハプテン(16)の 添加あるいは各種人工コファクター分子を反応系から除くことに よって完全に阻害された。このことより、本抗体が触媒する反応 は抗体の抗原結合部位で起こり、その触媒反応には人工コファク ター分子が必須であることが判明した。
6.3.β-脱離反応
前項で述べたアシル基転移反応の結果から、抗原結合部位に人 工コファクター分子結合部位を構築できたことが明らかとなっ た。また、ホロ酵素型触媒抗体の最大の特徴と利点は、人工コ ファクター分子を入れ替えることにより触媒反応の種類およびそ の触媒機構を制御できることである。そこで、人工コファクター 分子としてフェニル酢酸誘導体(24)を用い,塩基触媒によるβ- 脱離反応を検討した。その結果、抗体25E2がβ-ハロケトン (23)からエノン(25)へのβ-脱離反応を触媒することがわかっ た。一方、抗体27C1 はこのβ-脱離反応を触媒しなかった。詳 細な反応速度論的解析の結果から、Km24=122μM,Km23=864μM,kcat=0.89min-1と決定した。またkcatlKm23/lkuncat=2.4×105と非常に大きな速度加速を示した。
6.4.アルドール反応および脱炭酸反応
アルドール反応は有機合成化学のみならず、生物学においてもC-C結合形成反応として最も重要で基本的な反応のひとつであ
る。天然においても、アルドール反応を触媒する多くの酵素が見いだされている。これらアルドラーゼはその触媒機構に基づき大
きく二つのクラスに分けることができる。その中でも、クラス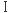 アルドラーゼはその活性部位にリジン残基をもち、アルドール供
与体である基質とのシッフ塩基の形成を経て、アルドール反応を触媒する。そこで、一級アミンコファクター(19)を用い、クラ
スアルドラーゼ型のアルドール反応の検討を行った。その結果、両抗体とも4.4×104倍の速度加速を示し、非常に効率的にアルドール反応を触媒することがわかった。また、エナミンの前駆体であるイミニウムイオン中間体を捕捉できたことにより、両抗体が触媒するアルドール反応はクラスアルドラーゼと同様にエナミン機構で反応が進行することが示唆された。
アルドラーゼはその活性部位にリジン残基をもち、アルドール供
与体である基質とのシッフ塩基の形成を経て、アルドール反応を触媒する。そこで、一級アミンコファクター(19)を用い、クラ
スアルドラーゼ型のアルドール反応の検討を行った。その結果、両抗体とも4.4×104倍の速度加速を示し、非常に効率的にアルドール反応を触媒することがわかった。また、エナミンの前駆体であるイミニウムイオン中間体を捕捉できたことにより、両抗体が触媒するアルドール反応はクラスアルドラーゼと同様にエナミン機構で反応が進行することが示唆された。
クラスアルドラーゼは同様のエナミン機構によりβ-ケト酸
の脱炭酸反応も触媒することが知られている。そこで、一級ア
ミンコファクター(19)を用い、クラスアルドラーゼ型のβ-ケ
ト酸(28)の脱炭酸反応を検討した(図11)。その結果、両抗体
とも非常に効率的に脱炭酸反応を触媒することがわかった。抗
体27C1の詳細な反応速度論的解析の結果から、Km19= 6.9mM,Km28=2.1mM,kcat=1.29min-1と決定した。また、kcatlKm 28lkuncat=1.4×105と非常に大きな速度加速を示すことがわかった。
ホロ酵素型触媒抗体の開発を目指して、ハプテンの設計および 合成、マウスへの免疫、モノクローナル抗体の作製、活性スクリー ニングを行い、触媒抗体25E2,27C1の獲得に成功した。また、 これら抗体は人工コファクター分子を入れ替えることによりアシ ル基転移反応、β-脱離反応、アルドール反応および脱炭酸反応 と触媒反応の種類および触媒機構を制御可能であり、さらに、こ れまで作製されてきた類似反応あるいは同様の反応を触媒する触 媒抗体と比較しても同程度あるいはそれ以上の触媒活性をもつこ とがわかった。また、このように一つの生体触媒が触媒機構の異 なる様々な反応を効率よく触媒する例は触媒抗体においてはもち ろんのこと天然酵素にも例がなく非常に興味深い。
7.おわりに:酵素と抗体
酵素は、多様な分子の発生と適応による選択を繰り返し、長い 年月をかけて進化してきた。まず、エクソン・シャフリングや遺 伝子重複によって多様性が生まれ、初生の酵素が選択される。そ の後、突然変異の蓄積にともなってさらに多様性を増やし、より 適応した活性をもつ酵素が選択される。このように酵素の分子進 化は多様性の発生と触媒効率に基づく選択によって起こるわけだ が、免疫システムの抗体タンパク質の産生機構と比較した時、図12に示すように多くの類似点を見いだすことができる。すなわ ち、免疫システムも酵素と同じ分子進化の仕組みを使って、目的 とした抗原結合性の抗体タンパク質を産生する。抗体にしろ、酵 素にしろ、分子進化の原動力は多様性の発生と選択である。しか も、免疫システムの場合、2~3週間で抗体タンパク質が産生す るので、免疫システム自身を進化分子工学の手法として利用する ことができる。
抗体酵素とは生体のもつ免疫システムを利用して、多種多様な 抗体タンパク質のレパートリーの中から、ハプテンのデザインに より望んだ触媒機能を持つ抗体を選別しようとするものである。 これは、酵素が何億年という長い年月をかけて進化を繰り返し選 別されることによって、高度な機能を獲得してきたのと似ており、 機能デザインのための最も合理的な方法とも考えられる。1986年に最初の抗体酵素が見いだされてから、今日までに多くの抗体 酵素が作製され、ペリ環状反応から加水分解反応までのいろいろ な化学反応が抗体によって触媒されることがわかってきた。現在 の物質生産プロセスは化学法や酵素法によって行われているが、 その幾つかは抗体酵素で置き換えることが可能であろう。近い将 来、第3 番目の方法として抗体酵素法が確立されることを期待し ている。

| 筆者紹介 | |
|---|---|
| 氏名 | 藤井 郁雄 (Ikuo Fujii) |
| 連絡先 | 大阪府立大学大学院理学系研究科 生物科学専攻生命化学研究室・教授 〒599-8570 大阪府堺市中区学園町1-1 Tel: 096-373-5313 Fax: 096-373-5314 e-mail:fujii@b.s.osakafu-u.ac.jp HP:http://if.b.s.osakafu-u.ac.jp/ |
| 略歴 | 1986年 九州大学院薬学研究科博士課程修了(薬学博士) 1986年 九州大学薬学部助手 1988年 ロックフェラー大学(米国)博士研究員 1989-1991年 スクリプス研究所 (米国) にて、博士研究員として触媒抗体の開発に従事 1991-2003年 タンパク工学研究所および生物分子工学研究所にて抗体工学に従事 2003年-4月より現職 |
| 進化分子工学を基盤とする新規生体機能分子の設計と創出 | |
| Copyright(c) 1996-2010 DOJINDO LABORATORIES,ALL Rights Reserved. |