乮姅乯摨恗壔妛尋媶強丂嵅乆杮丂堦旤
丂嫸媿昦乮BSE乯偺僷僯僢僋偑擔杮傪廝偭偰偄傞偑丄杮応僀僊儕僗偱傕BSE偐傜偺姶愼偲峫偊傜傟傞曄堎宆僋儘僀僣僼僃儖僩丒儎僐僽昦乮vCJD乯偺姵幰悢偼婛偵110柤傪墇偊丄埶慠偲偟偰憹壛偟偰偄傞丅丂偙傟傑偱丄媿偺昦婥偱恖傊偺姶愼偼側偄偲峫偊傜傟偰偄偨BSE偩偑丄1996擭3寧丄僀僊儕僗寬峃曐尟徣偑岞幃偵姶愼傪擣傔偨偺偑崱擔偺僷僯僢僋偺敪抂偲側偭偰偄傞丅丂椙偔抦傜傟偰偄傞傛偆偵丄尨場偲側傞僾儕僆儞抈敀幙偼惓忢擼慻怐偵傕敪尰偟偰偍傝乮惓忢僾儕僆儞PrPC乯丄偙傟偑壗傜偐偺嶌梡偱 b-僔乕僩峔憿偵晉傫偩傾僀僜僼僅乕儉偵堏峴偟乮堎忢僾儕僆儞PrP Sc乯丄擼撪偵嬅廤偡傞偙偲偱恄宱幘姵傪堷偒婲偙偡偲峫偊傜傟偰偄傞丅丂嵟戝偺撲偼丄PrPC偐傜PrPSc傊偺2師峔憿偺曄姺偑偳偺傛偆偵婲偙傞偐偱偁傞偑丄傾儈儘僀僪乕僔僗摨條丄徻嵶側婡峔偼暘偐偭偰偄側偄丅丂b-僔乕僩抈敀幙偑帺屓怗攠揑偵嬅廤偟偰偄偔偺偼懠偺傾儈儘僀僪抈敀幙偲摨偠偩偑丄戝偒側堘偄偼僾儕僆儞抈敀幙偑姶愼偡傞偙偲偱偁傞丅丂惗柦懱偲偼尵偊側偄扨側傞暔幙偑姶愼丒揱攄偟偰偄偔晄巚媍偝偼丄懡偔偺壢妛幰偺娭怱傪堷偄偰偄傞丅
丂
尰嵼PrPSc偺専弌偼丄擼慻怐傪儂儌僕僱乕僩偟丄僾儘僥傾乕僛偱張棟偟摼傜傟傞俶枛抂懁偑彍嫀偝傟偨142傾儈僲巁偐傜側傞僐傾晹暘乮PrP27-30丄恾1乯傪丄峈懱傪梡偄偰専弌偡傞曽朄偱峴傢傟偰偄傞乮BSE偺応崌丄ELISA偵傛傞堦師僗僋儕乕僯儞僌丄峏偵 Western blot媦傃慻怐柶塽愼怓偵傛傞妋掕恌抐偑峴傢傟偰偄傞乯丅丂vCJD偺応崌偼庒擭偱敪徢偟偰偍傝丄偟偐傕丄敪徢偟偰偐傜偺恑峴偑憗偄丅丂PrPSc偼丄椺奜揑偵梤偺儕儞僷慻怐偱専弌偝傟傞偑丄偦傟埲奜丄BSE傗vCJD偱偼愽暁婜偺惗慜恌抐偼晄壜擻偱偁傞丅
丂柍榑丄寣塼側偳傪梡偄傞惗慜恌抐傪壜擻偵偡傞峈懱偺奐敪偼悽奅拞偱嫞偭偰峴傢傟偰偄傞偑丄偦偺偨傔偵偼丄寣拞側偳偺旝検 PrPSc傪専弌偱偒傞姶搙偑昁梫偱偁傞丅丂栜榑偦傟偼丄扨偵尰嵼偺峈懱専弌偺姶搙偱偼専弌偱偒側偄偩偗偱丄嬌旝検偼懚嵼偟偰偄傞偲偄偆慜採偱偁傞丅丂偦偆偱偁傟偽丄PCR偺傛偆偵恖岺揑偵PrP Sc傪憹暆偱偒傟偽夝寛偡傞丅丂嬐偐偺PrPSc 偑妀偲側偭偰丄懱撪偱師乆偵PrPC傪PrP Sc偵曄偊偰偄偔姶愼偺僾儘僙僗偲婎杮揑偵摨偠偱偁傞丅
丂
恾1丂僾儕僆儞抈敀幙偺2師峔憿乮209傾儈僲巁乯
丂
丂Saborio傜1)偼protein-misfolding cyclic amplification乮PMCA乯偲屇偽傟傞憹暆僔僗僥儉傪峫埬偟偨乮恾2乯丅丂僗僋儗僀僺乕姶愼僴儉僗僞乕偺擼儂儌僕僱乕僩傪PrPSc偑専弌偱偒側偔側傞傑偱婓庍偟丄偦傟傪惓忢偺僴儉僗僞乕偺擼儂儌僕僱乕僩偲37亷偱僀儞僉儏儀乕僩偡傞偲PrPSc偑専弌偝傟偰偔傞乮proteinase K 偱張棟偟丄PrP27-30 傪専弌乯丅丂奅柺妶惈嵻偺懚嵼壓偱挻壒攇張棟傪峴偆偲嬅廤懱傪暘嶶偝偣傞偙偲偑偱偒傞偺偱丄峏偵摨條偺僒僀僋儖偱憹暆偡傞偙偲偑壜擻偱偁傞丅丂尨棟偼娙扨偩偑丄廳梫側億僀儞僩偼PrPC偺嫙媼尮偲偟偰擼儂儌僕僱乕僩傪梡偄偰偄傞偙偲偱偁傞丅丂惛惢偟偨PrPC傪PrPSc偲愙怗偝偣傞偙偲偱丄PrP C仺PrPSc偺曄姺偑婲偙傞偙偲偼抦傜傟偰偄偨偑 2)丄偦偺宯偱偼岠棪偼旕忢偵埆偔憹暆偡傞偙偲偼偱偒側偄丅丂偮傑傝丄嬅廤斀墳傪懀恑偡傞壗傜偐偺場巕乮椺偊偽 Protein X3)乯偺懚嵼偑昁恵偱丄儂儌僕僱乕僩傪梡偄傞昁梫偑偁傞丅丂PrP C偐傜PrPSc傊偺儕僼僅乕儖僨傿儞僌傪怗攠偡傞僔儍儁儘儞暘巕偺懚嵼偑媈傢傟偰偄傞偑丄偦傟偼PrP Sc帺恎偐傕抦傟側偄丅
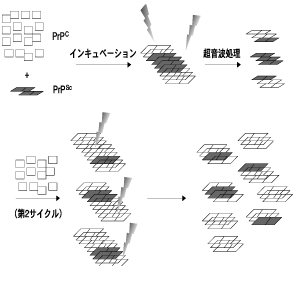
恾2丂PMCA僒僀僋儖
曬崘偝傟偨専弌姶搙偼丄10,000 攞埲忋偺婓庍偱丄10夞偺PMCA僒僀僋儖傪峴偄丄6乣12 pg乮0.2乣0.4 amol乯偺PrPSc偺専弌偑壜擻偱偁傞丅丂PrP Sc偲偄偆僥儞僾儗乕僩柍偟偱偼丄in vitro偱PrPSc傪嶌傝弌偡偙偲偑偱偒側偄側偳丄僾儕僆儞昦偺撲偼傑偩懡偄偑丄彮側偔偲傕in vivo偱偺尰徾傪棙梡偟偨崱夞偺媄弍偼丄僾儕僆儞昦偺憗婜恌抐偵峷專偡傞偙偲偼妋偐偱偁傞丅
丂
嶲峫暥專
1) G. P. Saborio, B. Permanne and C. Soto, Nature, 411, 810 (2001).
2) D. A. Kocisko, J. H. Come, S. A. Priola, B. Chesebro, G. J. Raymond,
P. T. Lansbury amd B. Caughey, Nature, 370, 471 (1994).
3) S. B. Prusiner, Proc. Natl. Acad. Sci., 95, 13363 (1998).