 |
(株)同仁化学研究所 鍋島哲二
|
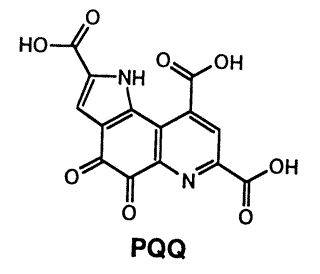
PQQ (pyrroloquinoline quinone) は次のような2段階の2電子酸化還元挙動をとるユニークな化合物である。
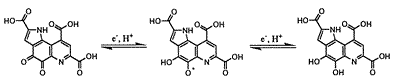
Fig. 1 PQQの酸化還元反応
1964年、Haugeらはグルコース脱水素酵素(GDH)中にNADH やフラビンとは異なる新たな補酵素が含まれることを報告した 1)。これが、PQQに関する最初の報告といわれている。1979年にこ の補酵素のX線解析がなされ、その構造が決定された 2)。NADH、フラビンに次ぐ「第3の補酵素」といわれるPQQの発見である。
PQQの研究は初期にはDuineらのグループによって精力的に 研究がなされ、GDH3) 、メタノール脱水素酵素(MDH)4)をはじめ 様々な脱水素酵素、酸化酵素の活性中心となっていることが明ら かとなった5)。当時は、グラム陰性細菌などの微生物にしか存在し ない特殊な酵素に含まれるものと思われていたが、その後、哺乳 類のアミン酸化酵素にも存在することが明らかとなるなど 6)、広く一般的な生物に存在していることが分かった。
このような、PQQを活性中心とする一連の酵素のなかの一つに グルコース脱水素酵素(GDH)がある。GDHはグルコースの1 位の水酸基を脱水素化しグルコノ-δ-ラクトンを生成するため、 糖尿病のスクリーニング用のセンサーとして用いる研究が進めら れている。同様な機能をもつ酵素にグルコース酸化酵素(GOD)が あり、酵素法を用いたグルコースの測定には酵素の安定性といっ た扱いやすさの面からGODを用いたセンサーが汎用されている。 しかしながら、このセンサーは溶存酸素に影響されるという問題点を もっている。
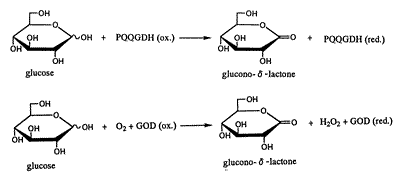
Fig. 2 GDHとGODによるグルコースの酸化
1984年にCassらはGODを用いてフェロセンをメディエー ターとするグルコースセンサーを報告し7)、その2年後にD'Costa らはGDHを用いて、Cassらが作製したものと同様なフェロセン をメディエーターとする電極を作製して比較した。その結果、 GODを用いたセンサーのレスポンスが60~90秒であるのに対 しGDHの場合では10~20秒と速く、かつ溶存酸素に影響され ない利点をもっていることが示された8)。グルコースセンサーとし ては正確さ、速度の面から有利なGDHであるが、酵素が熱的に不 安定だという問題点がある。早出らはGDHのアミノ酸配列を変 異させることで熱安定性を増す検討を行っている9,10)
PQQはセンサーとしての利用のほかに、酢酸を醸造する際に発
酵の誘導期間を短縮するなど、微生物の生育に関して有用である
ことが知られていたが11,12)、哺乳類など高等生物の生理活性物質
としても興味深い現象が知られるようになった。1989年にPQQ
欠乏マウスが報告され、生理活性物質としてのPQQの研究が盛
んになった。Killgoreらは、PQQを食餌に含むマウスと含まない
マウスを比較し成長過程を調べたところPQQをほとんど含まな
い食餌で生育させたマウスでは正常の2/3程度しか体重の増加が
なく、皮膚のコラーゲンの架橋が少なくなるなど、生育に大きな
差があることを示した13)。
このことからPQQは哺乳類の必須栄養素であると考えられる
ようになった。
熊沢らは、PQQをフェニルトリメチルアンモニウム(PTMA)で 誘導体化し、GC/MSにより存在量を測定した14) 。その結果、ヒトの臓器では脾臓 (5.9±3.4 ng/g)や膵臓(4.1±3.6 ng/g)など、多い部分でも数ng/gであり、一方、食物ではパセリで34.2±11.6 ng/gとその一桁高い値である15)。このことから、哺乳類は食物よ りPQQを得ているのではないかと述べている。
PQQは他の生理活性についても様々な研究がなされている。ヘ ム鉄が、Fe(IV)、Fe(V)等の高酸化状態になることを防ぐことや 16)、活性酸素と反応することなど17,18) 、酸化状態の制御、ラジカルの生成を抑える等の働きをもつことが示されている。
Aizenmanらはラットの大脳皮質神経細胞のN- メチルアスパラギン酸(NMDA)レセプターをDTTで還元して、開いたCaチャン ネルがPQQの添加によって正常に戻ることを示した19) 。これはPQQが還元されたレセプターを酸化して保護する機能をもつ事を 示している。
PQQは、このようにラジカルスカベンジャーとしての生理活性 を有していると報告されている。一方で、Kawanishiらは銅イオ ン存在下で、PQQがDNAに損傷を与えることを報告している 20)。彼等はPQQとCu(II)、NADHの存在する系でDNAのシトシン とチミンが損傷を受けることを示した。バソクプロイン(Cu(I)と 特異的に反応する試薬)がある場合は、Cu(I)がマスキングされ DNAの損傷が起きない。これは、NADHがPQQにより酸化され る際に発生する活性酸素と、Cu(II)が発生したPQQラジカルに より還元されて生じるCu(I)が反応して、生成する活性種により DNAが損傷されると考えられている。
PQQの研究は、特異な酸化還元機構から酵素の活性中心として のみならず、生理活性物質としても興味深い一面をもっている。ま た最近、金基板に固定化し電子伝達のメディエーターとする電極 も報告される21)など、その利用はますます広がり、今後の研究が 期待される。
1) J.G.Hauge,J.Biol.Chem., 239,3630 (1964).
2) S.A.Salisbury,J.S.Forrest,W.B.T.Cruse,O.Kennard, Nature, 280,843(1979).
3) J.A.Duine,J.Frank,J.K.van Zeeland,FEBS Lett.,108,443 (1979).
4) J.A.Duine,J.Frank,P.E.J.Verwiel,Eur.J.Chem .,108,187 (1980).
5) M.Ameyama,K.Matsushita,Y.Ohno,E.Shinagawa,O. Adachi,FEBS Lett.,130,179(1981).
6) C.L.Lovenstein-Verbeek,J.A.Jongejan,J.Frank,J.A. Duine,FEBS Lett.,170,305(1984).
7) A.E.G.Cass,G.Davis,G.D.Francis,H.A.O.Hill,W.J.Aston, I.
J.Higgins,E.V.Plotkin,L.D.L.Scott,A.P.E.Turner,Anal. Chem.,56,667(1984).
8) E.J.D'Costa,I.J.Higgins,A.P.F.Turner,Biosensors ,2,71 (1986).
9) K.Sode,T.Ootera,M.Shirahane,AB Witarto,S.Igarashi,H. Yoshida,Enzyme Microb.Tecnol.,26,491(2000).
10) K.Sode,K.Ito,A.B.Witarto,K.Watanabe,H.Yoshida,P. Postma,J.Biotechnol.,49,239(1996).
11) 飴山 寛,現代化学,7月号, p.46(1985).
12) 飴山 寛,松田 泰樹,久保田 昭正,瀧本 晃一,足立 収生,日本醸造協会誌,82,587(1987).
13) J.Killgore,C.Smidt,L.Duich,N,R,Chapman,D,Tinker,K. Reiser,M.Melko,D.Hyde,R.B.Rucker,Science,
245,850 (1989).
14) T.Kumazawa,H.Seno,T.Urakami,T.Matsumoto.O. Suzuki,Biochim.Biophys.Acta,1156,62
(1992).
15) T.Kumazawa,K.Sato,H.Seno,A.Ishii,O.Suzuki, Biochem. J., 307,331(1995).
16) F.Xu,C.P.Mark,K.S.Quandt,M.Schlafer,V.Massey,D.E. Hultquist,Biochem.Biophys.Res.Commun.,193
,434 (1993).
17) P.M.Gallop,M.A.Paz,R.Fluckiger,E.Henson,Connect. Tissue Res.,29,153(1993).
18) S.Itoh,Y.Ohshiro,T.Agawa,Bull.Chem.Soc.Jpn ., 59, 1911(1986).
19) E.Aizenman,F.E.Jensen,P.M.Gallop,P.A.Rosenberg,L. H.Tang, Neurosci.Lett.,168.189(1994).
20) Y.Hikaru,S.Kawanishi,FEBS Lett.,393,317(1996).
21) E.Katz,V.Heleg-Shabtai,A.Bardea,I.Willner,H,K.Rau,W. Haehnel,Biosens.Bioelectron.,13,741(1998).