僄僀僘偐傜尒偨姶愼徢尋媶偺嵟慜慄
偦偺4丂HIV姶愼偲廻庡場巕
孎杮戝妛戝妛堾堛妛栻妛尋媶晹姶愼杊屼暘栰丂慜揷梞彆
|
1.丂偼偠傔偵
丂僂僀儖僗棻巕偵偼堚揱忣曬偲偟偰偺妀巁偲偦傟傪曐岇偡傞帀幙枌傗彮悢偺峺慺側偳偺抈敀幙偑懚嵼偟偰偄傞偩偗偱丄偦傟帺恎偺傒偱憹怋偡傞偙偲偼偱偒側偄丅僂僀儖僗暋惢偺偨傔偵偼嵶朎撪偵怤擖偟偰丄嵶朎偵懚嵼偟偰偄傞庬乆偺峺慺側偳偺廻庡桼棃偺惉暘傪棙梡偡傞昁梫偑偁傝丄偟偨偑偭偰嵶朎偵偦偺傛偆側惉暘偑旛傢偭偰偄側偄応崌偼巕懛僂僀儖僗傪嶻惗偱偒側偄丅堦斒偵丄偁傞摿掕偺僂僀儖僗偺庬摿堎惈傗嵶朎巜岦惈偼偙偺傛偆側僂僀儖僗暋惢偵娭楢偡傞庬乆偺僂僀儖僗暋惢曗彆場巕偺桳柍偵傛傝寛掕偝傟偰偄傞丅HIV傪娷傔偨儗僩儘僂僀儖僗傕摨條偵庬乆偺廻庡場巕偺桳柍偑偦偺庬摿堎惈傗嵶朎巜岦惈傪寛掕偟偰偄傞偙偲偑柧傜偐偲
側偭偰偄傞偑丄嵟嬤丄儗僩儘僂僀儖僗偺憹怋傪惂屼偟偰偄傞廻庡場巕偲偟偰丄偦偺憹怋傪愊嬌揑偵梷偊傞傛偆側場巕偑摨掕偝傟丄偙偺傛偆側儗僩儘僂僀儖僗姶愼掞峈惈場巕偑偦偺庬摿堎惈傪寛掕偟偰偄傞偙偲偑柧傜偐偲側偭偰偒偨丅杮峞偱偼偙偺傛偆側姶愼掞峈惈場巕偲偟偰偺廻庡場巕偵徟揰傪峣偭偰夝愢偡傞丅
仯儁乕僕偺僩僢僾傊
2. TRIM5兛
丂HIV偺婲尮偼傾僼儕僇偵惗懅偟偰偄傞傾僼儕僇儈僪儕僓儖傗傾僇僎僓儖側偳偺媽悽奅僓儖偵懚嵼偟偰偄傞僒儖柶塽晄慡僂僀儖僗(SIV)偱偁傞偲峫偊傜傟偰偄傞偑丄SIV偐傜恑壔偟偨偲峫偊傜傟傞HIV偼僒儖桼棃嵶朎偱偼憹怋偱偒側偄偙偲偑埲慜傛傝曬崘偝傟偰偄偨丅偙偺傛偆側庬摿堎惈偼堦斒偵偦偺僂僀儖僗偺庴梕懱暘巕偺堘偄偵傛偭偰愢柧偝傟傞偙偲偑傛偔抦傜傟偰偄傞丅偟偐偟僒儖偺CD4暘巕傕HIV偺庴梕懱偱偁傞僸僩CD4暘巕偲摨條偵HIV偺庴梕懱偲偟偰婡擻偡傞偙偲偑傢偐傝丄偝傜偵HIV偺僂僀儖僗怤擖偺岠棪傪忋偘偰傕HIV偺嵶朎撪暋惢偑慾奞偝傟偰偄傞偙偲偑敾柧偟偨丅偡側傢偪HIV偺応崌丄僒儖嵶朎怤擖屻媡揮幨埲慜偺儗儀儖偱偦偺暋惢偑慾奞偝傟偰偄偨偺偱偁傞丅偙偺傛偆側僂僀儖僗暋惢偺掆巭偼堦斒揑偵偼慜弎偺傛偆偵壗傜偐偺僂僀儖僗暋惢偵昁恵側嵶朎撪偵懚嵼偡傞廻庡場巕偑寚擛偟偰偄傞偙偲偱愢柧偝傟傞偑丄戝検偺僂僀儖僗棻巕偱嵶朎傪慜張棟偟偰偍偔偲姶愼偑惉棫偡傞傛偆偵側傞偙偲偐傜丄僂僀儖僗抈敀幙偵傛傝朞榓偝傟傞姶愼掞峈惈場巕偺娭梌偑帵嵈偝傟偨丅偦偙偱Sodroski傜偼丄傾僇僎僓儖桼棃cDNA儔僀僽儔儕乕傪僸僩嵶朎偵摫擖偟HIV掞峈惈傪晅梌偡傞堚揱巕傪摨掕偟偨丅摨掕偟偨暘巕偼RING finger, B-box, Coiled-coil偲屇偽傟傞3偮偺僪儊僀儞傪桳偡傞Tripartite僼傽儈儕乕 偵懏偟偰偍傝丄偝傜偵偦偺C枛抂偵SPRY (B30.2) 偲屇偽傟傞僪儊僀儞峔憿傪桳偡傞暘巕丄TRIM5兛偱偁偭偨乮恾乯丅僒儖TRIM5兛偼HIV偺姶愼傪慾奞偡傞偙偲偑偱偒傞偑丄僸僩TRIM5兛偼HIV偺姶愼傪慾奞偡傞偙偲偑偱偒偢丄庬摿堎惈傪愢柧偱偒傞暘巕偱偁傞偙偲偑帵偝傟偨丅偝傜偵庬摿堎揑姶愼掞峈惈偼偦傟偧傟偺庬偺SPRY僪儊僀儞偑寛掕偟偰偄傞偙偲偑柧傜偐偲側偭偨丅
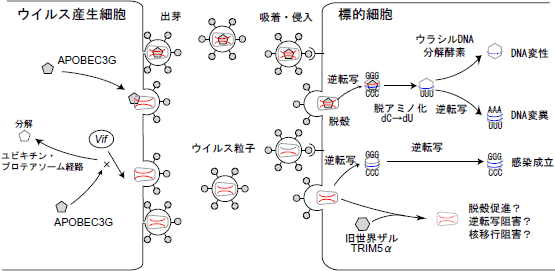 |
乮恾乯HIV-1偺姶愼掞峈惈場巕APOBEC3G丒TRIM5
僂僀儖僗嶻惗嵶朎偵Vif抈敀幙偑懚嵼偟偰偄傞偲APOBEC3G偼儐價僉僠儞丒僾儘僥傾僜乕儉宱楬偵傛偭偰暘夝偝傟傞偑丄Vif抈敀幙偑懚嵼偟側偄偲僂僀儖僗棻巕偵APOBEC3G偑庢傝崬傑傟傞丅昗揑嵶朎撪偵僂僀儖僗偑怤擖丒扙妅偟偨屻丄僂僀儖僗RNA乮愒乯偐傜媡揮幨偝傟偨儅僀僫僗嵔僂僀儖僗DNA乮惵乯偺僔僩僔儞巆婎偑APOBEC3G偺僔僠僕儞僨傾儈僫乕僛偺妶惈偵傛傝僂儔僔儖巆婎傊偺曄姺傪庴偗傞丅偦偺寢壥僾儔僗嵔僂僀儖僗DNA乮惵乯偵僌傾僯儞偐傜傾僨僯儞傊偺曄堎偑摫擖偝傟丄傾儈僲巁偺曄堎傗掆巭僐僪儞偺弌尰偵傛傝僂僀儖僗抈敀幙崌惉偑慾奞偝傟傞丅傑偨丄僂儔僔儖偵曄姺偝傟偨DNA偼僂儔僔儖DNA暘夝峺慺偵傛傝攋夡偝傟丄椉幰偺嶌梡偵傛傝僂僀儖僗偺暋惢偑慾奞偝傟傞丅堦曽丄媽悽奅僓儖TRIM5兛偼僂僀儖僗怤擖屻丄僂僀儖僗僇僾僔僪抈敀幙偲斀墳偟丄扙妅丒媡揮幨丒妀堏峴傪慾奞偟偰偄傞偲峫偊傜傟偰偄傞丅 |
丂堦曽丄埲慜傛傝僇僾僔僪抈敀幙偲僒僀僋儘僼傿儕儞A偺寢崌傪慾奞偡傞暘巕偱偁傞僒僀僋儘僗億儕儞偑HIV-1偺姶愼傪慾奞偡傞偙偲偑傢偐偭偰偄偨丅偙傟偼昗揑嵶朎偵懚嵼偡傞姶愼掞峈惈場巕偐傜摝旔偡傞偨傔偵僂僀儖僗偑偦偺僇僾僔僪抈敀幙偺棫懱峔憿傪僒僀僋儘僼傿儕儞A偲偺寢崌偵傛傝曄壔偝偣偰偄傞偑丄僒僀僋儘僗億儕儞偼偦偺棫懱峔憿傪姶愼掞峈惈場巕偵斀墳偱偒傞宍偵栠偡偙偲偵傛偭偰姶愼傪慾奞偟偰偄傞偲峫偊傜傟偰偄傞丅嬃偄偨偙偲偵丄堦斒揑偵偼怴悽奅僓儖偼HIV-1偵懳偟偰姶愼姶庴惈偱偁傞偑丄怴悽奅僓儖偺堦庬偱偁傞儓僓儖偱偼HIV-1姶愼偵掞峈惈偱偁傞偙偲偑帵偝傟丄偝傜偵僒僀僋儘僗億儕儞揧壛偵傛傝姶愼姶庴惈偑忋
徃偟偰偔傞偙偲偑傢偐偭偨丅偙傟偼儓僓儖偱偼TRIM5兛偺SPRY僪儊僀儞偺拞偵丄儗僩儘僩儔儞僗億僝儞偵傛偭偰僒僀僋儘僼傿儕儞A堚揱巕偑帩偪崬傑傟偨寢壥丄Tripartite儌僠乕僼偲僒僀僋儘僼傿儕儞A偺梈崌抈敀幙(TRIM-Cyp)偑敪尰偟偰偍傝丄HIV-1偺僇僾僔僪抈敀幙偲偙偺梈崌抈敀幙偑捈愙寢崌偡傞偙偲偑峈僂僀儖僗妶惈偵廳梫偱偁傞偙偲偑傢偐偭偨丅摨條偵傾僇僎僓儖TRIM5兛傕SPRY僪儊僀儞偲HIV-1偺僇僾僔僪抈敀幙偑憡屳嶌梡偡傞偙偲偑峈僂僀儖僗妶惈偵廳梫偱偁傞偲憐掕偝傟偰偄傞偑丄捈愙揑側寢崌偼妋擣偝傟偰偍傜偢丄偦偺姶愼慾奞婡峔偺徻嵶偵偮偄偰偼尰帪揰偱偼傛偔傢偐偭偰偄側偄丅傑偨HIV-1僇僾僔僪抈敀幙偺堦晹傪僒僀僋儘僼傿儕儞寢崌擻偺側偄SIV僇僾僔僪抈敀幙偵抲姺偡傞偲僒儖桼棃TRIM5兛偵傛傞HIV-1姶愼慾奞傪夞旔偡傞偙偲偑偱偒傞傛偆偵側傞偙偲偐傜丄僒僀僋儘僗億儕儞偵娭楢偡傞姶愼掞峈惈場
巕偲偟偰僸僩TRIM5兛偑峫偊傜傟偨偑丄尰帪揰偱偼斲掕揑側曬崘偑懡偄丅TRIM5兛埲奜偺姶愼掞峈惈場巕偑娭梌偟偰偄傞偺偐丄TRIM5兛偲憡屳嶌梡偡傞暘巕偑娭梌偟偰偄傞偺偐偼尰帪揰偱偼傢偐傜側偄丅傑偨嵟嬤丄僒儖TRIM5兛偼HIV-1偺扙妅傪堎忢偵懀恑偟偰偄傞偲偺曬崘偑偁傝丄扙妅偲偄偆尰嵼傛偔傢偐偭偰偄側偄暋惢夁掱傪抦傞偒偭偐偗偲側傞偐傕偟傟側偄丅傑偨丄僸僩偺TRIM5兛偼HIV-1偵懳偟偰偼傎偲傫偳姶愼慾奞岠壥偑側偄傕偺偺丄儅僂僗儗僩儘僂僀儖僗偺堦晹偵懳偟偰偼姶愼慾奞岠壥偑偁傝丄恑壔忋壗傜偐偺儗僩儘僂僀儖僗偺怤擖傪杊偖偨傔偵昁梫側場巕偱偁偭偨偺偱偼側偄偐偲悇應偝傟偰偄傞丅
仯儁乕僕偺僩僢僾傊
3. APOBEC3G
丂HIV-1偵偼vif偲屇偽傟傞傾僋僙僒儕乕堚揱巕偑懚嵼偟偰偄傞偑丄偙偺堚揱巕偺栶妱偵偮偄偰偼傛偔傢偐偭偰偄側偐偭偨丅vif堚揱巕寚懝僂僀儖僗偱傕憹怋偑壜擻側嵶朎偲憹怋偱偒側偄嵶朎偑偁傝丄憹怋偱偒側偄嵶朎偱偼僂僀儖僗怤擖屻媡揮幨偺儗儀儖偱慾奞偑偐偐偭偰偄傞偙偲偑曬崘偝傟偰偄偨丅傑偨偙傟傜偺姶愼旕姶庴惈嵶朎偲姶愼姶庴嵶朎傪嵶朎梈崌偝偣偰僿僥儘僇儕僆儞傪嶌惢偟偰偦偺宍幙傪傒偰傒傞偲丄姶愼旕姶庴惈嵶朎撪偵懚嵼偟偰偄傞壗傜偐偺廻庡場巕偑vif堚揱巕寚懝僂僀儖僗偵懳偟偰姶愼掞峈惈傪晅梌偟偰偄傞偙偲偑傢偐偭偨丅偦偙偱Malim傜偼姶愼旕姶庴惈嵶朎偲姶愼姶庴惈嵶朎偺敪尰傪斾妑偟偰丄姶愼旕姶庴惈嵶朎偵偺傒敪尰偟偰偄傞場巕傪摨掕偟偨偲偙傠丄偙偺場巕偼妀巁偺僔僩僔儞巆婎傪扙傾儈僲壔偟偰僂儔僔儖巆婎偵曄姺偡傞APOBEC3G偲偄偆僔僠僕儞僨傾儈僫乕僛偲偄偆峺慺偩偭偨丅APOBEC3G偼vif堚揱巕寚懝僂僀儖僗偱偼僂僀儖僗棻巕撪偵庢傝崬傑傟丄昗揑嵶朎偱僂僀儖僗RNA偐傜媡揮幨偝傟偨儅僀僫僗嵔DNA偺僔僩僔儞傪扙傾儈僲壔偟偰僂儔僔儖偵曄姺偡傞偙偲偵傛傝丄寢壥揑偵僾儔僗嵔DNA偵懡悢偺僌傾僯儞偐傜傾僨僯儞傊偺曄堎傪摫擖偟丄傾儈僲巁偺曄堎傗掆巭僐僪儞偺弌尰偵傛傝僂僀儖僗抈敀幙崌惉傪慾奞偡傞乮恾乯丅傑偨摨帪偵DNA嵔偵庢傝崬傑傟偨僂儔僔儖DNA偼僂儔僔儖DNA暘夝峺慺偵傛傝暘夝偝傟傞偲峫偊傜傟偰偄傞乮恾乯丅Vif抈敀幙偼僂僀儖僗嶻惗嵶朎撪偱APOBEC3G偲寢崌偡傞偙偲偵傛傝儐價僉僠儞丒僾儘僥傾僜乕儉宯偱APOBEC3G暘夝傪懀恑偟丄寢壥偲偟偰APOBEC3G偺棻巕撪庢傝崬傒傪慾奞偟丄僂僀儖僗偼嵶朎撪偱曄堎傪婲偙偡偙偲側偔憹怋偱偒傞傛偆偵側傞乮恾乯丅偟偐偟側偑傜APOBEC3G偺妶惈拞怱偵曄堎傪摫擖偟偰僔僠僕儞僨傾儈僫乕僛偺妶惈傪幐妶偝偣偰傕姶愼慾奞偑偐偐傞偲偺曬崘傗丄昗揑嵶朎偵懚嵼偟偰偄傞APOBEC3G偑Vif旕埶懚揑偵僂僀儖僗暋惢傪慾奞偟偰偄傞曬崘傕偁傝丄偦偺姶愼慾奞儊僇僯僘儉偵偮偄偰偼傑偩晄柧側揰傕懡偄丅傑偨APOBEC3G埲奜偵傕APOBEC3F傗APOBEC3B側偳懠偺僸僩APOBEC僼傽儈儕乕傗丄儔僢僩丒儅僂僗側偳偺APOBEC僼傽儈儕乕傕HIV-1偺姶愼傪慾奞偡傞偙偲偑師乆偲曬崘偝傟偨丅傑偨儅僂僗儗僩儘僂僀儖僗側偳懠庬偺儗僩儘僂僀儖僗姶愼傕僸僩APOBEC3G偵傛傝慾奞偱偒傞側偳丄惗暔庬偲儗僩儘僂僀儖僗偺恑壔偲偄偆娤揰傊偲奼偑傝傪尒偣偰偄傞丅嫽枴怺偄偙偲偼HIV-1偺Vif抈敀幙偼傾僼儕僇儈僪儕僓儖APOBEC3G傪拞榓偡傞偙偲偑偱偒側偄丅媡偵傾僼儕僇儈僪儕僓儖桼棃僒儖柶塽晄慡僂僀儖僗(SIVagm)偺Vif抈敀幙偼僸僩偺APOBEC3G傪拞榓偱偒偢丄偦偺庬摿堎惈偼APOBEC3G偺128斣栚偺傾儈僲巁偵傛傝寛掕偝傟偰偄偨丅偙傟偼APOBEC3G偑僶儕傾乕偲側傝懠偺惗暔庬偺儗僩儘僂僀儖僗怤擖傪杊偄偱偄傞偙偲傪堄枴偟偰偄傞偺偐傕偟傟側偄丅傑偨僂僀儖僗懁傕vif堚揱巕傪妉摼偡傞偙偲偵傛傝惗懱偺杊屼婡峔偐傜摝傟偰僂僀儖僗暋惢傪壜擻偵偟偰偄傞偲傕峫偊傜傟丄僂僀儖僗偲廻庡偺偟偺偓偁偄傪奯娫尒偰偄傞偺偐傕偟傟側偄丅
仯儁乕僕偺僩僢僾傊
4. 偍傢傝偵
丂TRIM5兛傗APOBEC3G偼忋弎偺傛偆偵懠庬偺儗僩儘僂僀儖僗偐傜偺姶愼傪杊屼偡傞栶妱傪桳偟偰偍傝丄尰嵼偱偼儗僩儘僂僀儖僗偵懳偡傞廻庡偺帺慠柶塽偺堦晹偲偟偰峫偊傜傟傞傛偆偵側偭偰偒偰偄傞丅崱屻偙偺傛偆側儗僩儘僂僀儖僗偺姶愼傪慾奞偡傞廻庡場巕偑怴偨偵暘棧偝傟傞偙偲偑梊憐偝傟丄偝傜偵偼儗僩儘僂僀儖僗埲奜偺僂僀儖僗偵懳偡傞姶愼掞峈惈場巕傕暘棧偝傟偰偔傞傕偺偲婜懸偝傟傞丅幚嵺偵APOBEC3G偼B宆娞墛僂僀儖僗偺丄TRIM僼傽儈儕乕偺堦晹偼僿儖儁僗僂僀儖僗傗僀儞僼儖僄儞僓僂僀儖僗側偳偺姶愼掞峈惈偵娭梌偟偰偄傞偙偲偑曬崘偝傟偰偍傝丄僂僀儖僗偵懳偡傞帺慠柶塽偺奣擮傪戝偒偔曄偊傞壜擻惈偑偁傞丅偟偐偟側偑傜奜棃惈偺僂僀儖僗偵懳偟偰偙傟傜偺暘巕傪廻庡偑杮棃偺栚揑偲偟偰旛偊偰偄傞傕偺側偺偐丄扨側傞嬼慠側偺偐偼媍榑偑懡偄丅嵟嬤APOBEC3G偑儗僩儘僩儔儞僗億僝儞傪梷惂偟偰偄傞偲偄偆曬崘偑偁傝丄僂僀儖僗偵尷傜偢嵶朎偵偲偭偰晄搒崌側僀儀儞僩傪梷惂偡傞偙偲偑杮棃偺栚揑側偺偐傕偟傟側偄丅傑偨偙偺傛偆側姶愼掞峈惈場巕偺嶌梡婡彉傪夝愅偡傞偙偲偵傛傝怴偨側峈HIV-1愴棯偑惗傑傟傞偙偲傪婜懸偟偨偄丅
| 挊幰徯夘 |
 |
巵柤丗慜揷 梞彆
強懏丗孎杮戝妛戝妛堾堛妛栻妛尋媶晹姶愼杊屼暘栰
廧強丗孎杮巗杮憫1-1-1
尋媶僥乕儅丗儗僩儘僂僀儖僗偺廻庡嵶朎傊偺怤擖婡峔偺夝柧
丂丂丂丂丂丂丂丂HIV偺峈HIV嵻偵懳偡傞懴惈妉摼婡峔偺夝柧
|
仯儁乕僕偺僩僢僾傊
|

